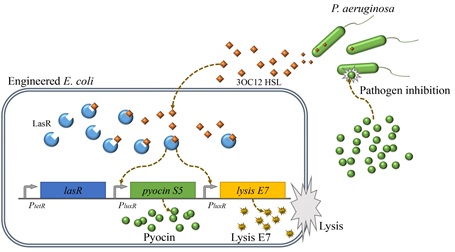
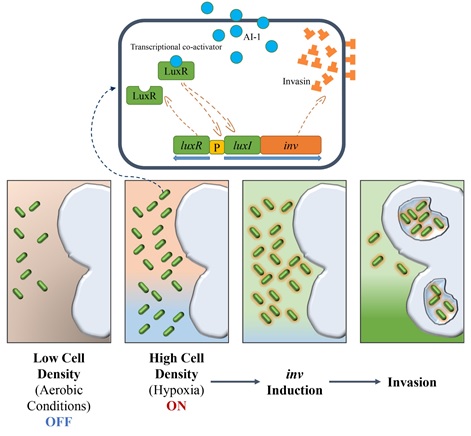
5. 고찰
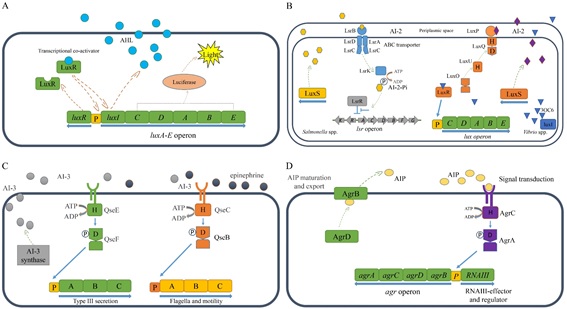
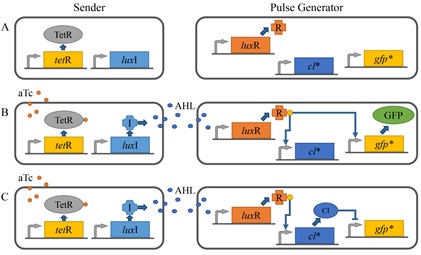
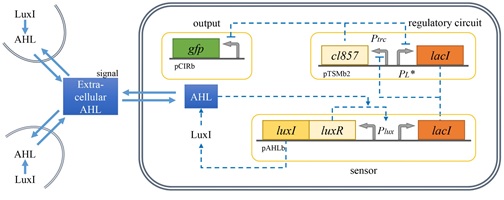
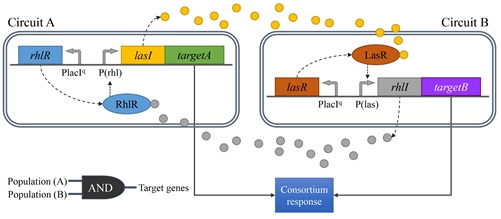
박테리아는 다양한 형태 커뮤니티의 구성원으로서 끊임없이 변화하는 외부 환경에 효율적으로 적응하기 위하여 신호전달물질, 센서, 신호전달 메커니즘을 포함하는 신호전달 시스템을 활용한다. 특히 AHL 이나 AIP 와 같은 신호전달 물질을 매개로 하는 쿼럼센싱 기반의 유전자 발현 조절기구는 다양한 형태의 커뮤니티를 형성하는 박테리아의 생존에 있어서 필수적인 세포간 신호전달 체계로 인식되고 있다. 따라서 여기서 언급한 대표적인 시스템들에 대한 연구 외에도 새로운 쿼럼센싱 신호전달계를 발굴하고 분석하기 위한 연구도 계속해서 활발히 진행되고 있다. 또한 최근에는 쿼럼센싱 시스템 정보를 활용하기 위한 연구도 부상하기 시작하였는데 신호전달 물질과 유사한 구조를 갖는 물질을 개발하기 위한 생명공학 연구가 그 예이다. 특히 이런 물질은 쿼럼센싱을 통해 병원성을 조절하는 세균을 제어하기 위한 새로운 항생제로 개발될 수 있는 가능성 때문에 주목 받고 있다. 아울러 세균의 쿼럼센싱 시스템 연구는 세포 내 혹은 세포간 신호전달, 혹은 동일 종 내 혹은 이종간 신호전달에 대한 정보와 나아가 다세포 생명체의 진화에 관한 정보를 제공할 수 있다는 점을 고려할 때 박테리아의 쿼럼센싱 시스템에 관한 정보와 모듈 소재는 합성생물학적으로 설계하고 제작하는 지능형 시스템의 효율적 운용을 위한 자가 조절 및 네트워크 조절 기구로 유용하게 활용될 수 있을 것이다.
6. 참고문헌
1. Ahmer BMM (2004) Cell-to cell signaling on Escherichia coli and Salmonella enterica. Mol Microbiol 52:933-945.
2. Anderson JC, Clarke EJ, Arkin AP, Voigt CA (2006) Environmentally controlled invasion of cancer cells by engineered bacteria. J Mol Biol 355:619–627.
3. Asad S, Opal SM (2008) Bench-to-bedside review: Quorum sensing and the role of cell-to-cell communication during invasive bacterial infection. Crit Care. 12:236.
4. Balagadde FK, Song H, Ozaki J, Collins CH, Barnet M, Arnold FH, Quake SR, You L (2008) A synthetic Escherichia coli predator–prey ecosystem. Mol Syst Biol 4:187.
5. Bassler BL (2002) Small talk. Cell-to-cell communication in bacteria. Cell 109:421-424.
6. Basu S, Mehreja R, Thiberge S, Chen MT, Weiss R (2004) Spatiotemporal control of gene expression with pulse-generating networks. Proc Natl Acad Sci USA 101:6355–6360.
7. Basu S, Gerchman Y, Collins CH, Arnold FH, Weiss R (2005) A synthetic multicellular system for programmed pattern formation. Nature 434:1130–1134.
8. Brenner K, Karig DK, Weiss R, Arnold FH (2007) Engineered bidirectional communication mediates a consensus in a microbial biofilm consortium. Proc Natl Acad Sci 104:17300–17304.
9. Garg N, Manchanda G, Kumar A (2014) Bacterial quorum sensing: circuits and applications. Antonie Van Leeuwenhoek. 105:289-305.
10. Gov Y, Borovok I, Korem M, Singh VK, Jayaswal RK, Wilkinson BJ, Rich SM, Balaban N (2004) Quorum sensing in staphylococci is regulated via phosphorylation of three conserved histidine residues. J Biol Chem 279:14665-14672.
11. Kobayashi H, Kaern M, Araki M, Chung K, Gardner TS, Cantor CR, Collins JJ (2004) Programmable cells: interfacing natural and engineered gene networks. Proc Natl Acad Sci USA 101:8414–8419.
12. Mangwani N, Dash HR, Chauhan A, Das S. (2012) Bacterial quorum sensing: functional features and potential applications in biotechnology. J Mol Microbiol Biotechnol. 22:215-227.
13. Nealson KH, Platt T, Hastings JW (1970) Cellular control of synthesis and activity of the bacterial luminescent system. J Bacteriol 104:313–322
14. Saeidi N, Wong CK, Lo TM, Nguyen HX, Ling H, Leong SSJ, Poh CL, Chang MW (2011) Engineering microbes to sense and eradicate Pseudomonas aeruginosa, a human pathogen. Mol Syst Biol 7:521.
15. Walters M, Sperandio V (2006) Autoinducer 3 and epinephrine signaling in the kinetics of locus of enterocyte effacement gene expression in enterohemorrhagic Escherichia coli. Infect Immun 74:5445-5455.
16. Waters CM, Bassler BL (2005) Quorum sensing: cell-to-cell communication in bacteria. Annu Rev Cell Dev Biol 21:319–346.
17. Williams P (2007) Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 153:3923–3938.
18. You L, Cox RS III, Weiss R, Arnold FH (2004) Programmed population control by cell–cell communication and regulated killing. Nature 428:868–871.









 박테리아 쿼럼센싱 시스템의 합성생물학적 응용
박테리아 쿼럼센싱 시스템의 합성생물학적 응용