방선균 이종숙주 생산시스템의 연구 동향
윤여준 교수
이화여자대학교
1. Introduction
자연계에 존재하는 수많은 미생물 중 방선균 (Streptomyces)은 대표적인 그램양성 (Gram-positive) 토양미생물로서 현재까지 보고된 항생제 중 약 60% 이상을 생산하며, 항생제 이외에도 면역억제제, 항암제, 항진균제, 제초제, 당뇨병치료제, 항기생충제 등의 다양한 생리활성물질을 생산하는 매우 매력적인 미생물이다. 그러나 새로운 생리활성물질을 발견하기 위한 다양한 노력에도 불구하고, 최근 몇 년 동안 방선균으로부터 새로운 생리활성물질의 발견은 감소하고 있다. 또한 이들 생리활성물질을 생산하는 야생 생산균주의 유전자 조작과 배양이 어려울 경우 및 유전자원의 실질적 활용을 위해서는 확보된 유전체 정보를 바탕으로 대량생산을 가능하게 하는 보편적인 기술의 개발이 필요하다. 고전적인 화학적 합성법이 아닌, 생합성 과정을 이해하고 이를 이용하여 구조변형 개량신약을 개발할 수 있는 ‘이종숙주 생산시스템’이 해결 방안이 될 수 있다. 특히 산업적으로 중요한 대표적 방선균 중 극히 일부만 유전자 조작이 가능하므로, 유용 생리활성물질 생합성 유전자를 유전자 조작이 용이한 이종숙주에 도입하고 발현하는 기술 개발이 필요하다 (그림 1).
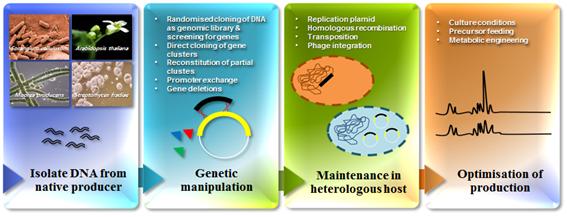
그림 1. 이종숙주 생산시스템의 개발
효과적인 이종숙주 생산시스템의 개발을 위해서는 안정된 발현벡터, 적합한 이종숙주의 선정, 생리활성물질 생합성 경로의 이해, 그리고 마지막으로 이종숙주 생산시스템의 안정적 발현을 위한 최적화된 방법의 고려가 필요하다. 이러한 이종숙주 생산시스템을 이용한 이차대사산물의 생산에 있어서 유전자 정보가 잘 알려져 있고, 전체 genome sequencing이 완료되어 있어, 대사체들이 잘 알려져 있는 방선균 Streptomyces coelicolor, S. lividans, S. albus J1074, S. avermitilis 등이 이차대사산물의 생산에 적합한 host로서 최적화된 발현 시스템을 갖추기에 많은 장점을 가지고 있다. 이러한 이유로 현재 많은 연구실과 산업 속에서 이종숙주로서 사용되고 있다.
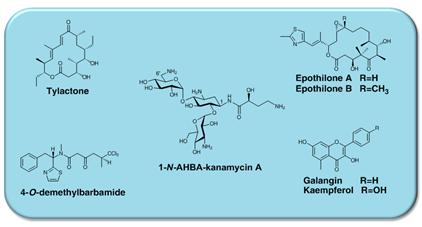
그림 2. 본 연구실에서 방선균 이종숙주 생산 시스템을 통하여 생산된 생리활성물질
2. 방선균에서 방선균으로의 이종숙주 발현
일반적인 cloning host로 S. albus J1074는 Salas group에서 처음 사용하여 fredericamycin의 이종숙주 발현에 사용되었다. S. griseus가 생산하는 항생제인 fredericamycin의 생합성 유전자집단을 이종숙주 S. albus J1074에서 high copy number plasmid를 사용하여 발현시켰을 때 120 mg/L가 생산되고 [1], single copy number φC31-integration으로 132 mg/L가 생산되었다 [2]. 반면에 동일한 fredericamycin pathway를 S. lividans에서 φC31 attB site를 이용하여 발현시켰을 때는 생산이 되지 않았다 [2].
S. coelicolor와 S. lividans는 genomic sequence가 두 방선균 모두 밝혀져 있으며 유전자조작 방법 역시 잘 알려져 있어 연구실에서 자주 사용되는 이종숙주 발현용 host이다. Novobiocin은 S. niveus에서 생산되는 항생물질로서 φC31 attB site를 이용하여 S. coelicolor M512와 S. lividans TK24에서 각각 이종숙주 발현 하였을 때 재미있는 결과를 나타내었다. 각각의 생산량이 31 mg/L와 1 mg/L로 현격한 차이를 보였다 [3]. 더 나아가 S. coelicolor M512에 이종숙주 발현을 하고, 배양 조건 optimize를 실시하였을 때 초기 생산량 31 mg/L에서 54 mg/L로 증가하고, 유전자 조작을 통하여 positive regulator 단백질을 암호화하고 있는 유전자를 과발현시켜 주었을 때 163 mg/L로 증가하는 결과를 나타내었다 [4].
본 연구팀은 이종숙주 생산시스템의 host로 S. venezuelae를 선정하고, pikromycin의 aglycon 생합성에 필요한 유전자들과 deoxysugar인 desosamine에 필요한 유전자들을 제거한 돌연변이주를 제작·개발하였다. 또한 거대생합성 유전자 발현에 적합한 E. coli-Streptomyces shuttle vector로서 동시 다중 발현이 가능한 replication vector와 integration vector를 고안하고, 외래 생합성 유전자집단의 효율적 안정적 발현을 위한 동시 다중 벡터 조합을 최적화 하였다. 이를 통하여 S. fradiae 유래 항생제인 tylosin의 약 50 kb에 달하는 거대 유전자 집단을 2종 동시 발현이 가능한 벡터 시스템에 tylosin polyketide synthase (Tyl PKS)를 나누어 클로닝한 후, PKS를 제거한 S. venezuelae 돌연변이주 내에서 발현하여 tylactone 0.5 mg/L과 desosaminyl tylactone을 성공적으로 생산하였다 [5].
또한, 결핵균과 폐렴균 등의 치료에 사용되어 온 가장 오래된 항생제 중 하나인 kanamycin을 생산하는 토양미생물 S. kanamyceticus는 유전자 조작이 거의 불가능해, kanamycin의 생합성 경로 규명은 전 세계 과학자들에게 풀리지 않는 숙제였다. 본 연구팀은 kanamycin 합성에 관여하는 모든 유전자를 선별한 후, 이들을 여러 개의 유전자 조각으로 잘라 레고 블록처럼 조립하여, 유전자 조작이 쉬운 이종숙주인 S. venezuelae에서 발현하고, 다양하게 조합된 유전자 세트에서 만들어낸 물질을 하나씩 확인하는 방법으로 kanamycin 생합성 경로를 밝혀내었다 [6].
3. 식물에서 방선균으로의 이종숙주 발현
식물 유래의 polyphenol 화합물은 항산화, 암예방, 항암, 항천식, 항염증, 항균 등의 다양한 생리활성을 갖으며, 그 중 flavonoid와 stilbene 화합물은 각종 기능성 식품, 화장품 및 의약품 개발의 잠재력이 있는 새로운 자원으로 중요한 연구 대상이다.
일본 동경대의 Horinouchi 그룹은 E. coli를 이종숙주로 사용하여 yeast인 Rhodotorula rubra의 PAL이 tyrosine을 기질로 사용하여 coumaric acid를 생산할 수 있다는 사실을 이용하여, 이를 방선균인 S. coelicolor A3(2)의 4-coumarate:CoA ligase (ScCCL), licorice plant인 Glycyrrhiza echinata의 CHS의 유전자들과 조합 발현하여 flavonoid 화합물 중 가장 간단한 구조의 flavanone인 naringenine을 소량 생산하는데 성공하였다 [7]. 또한 미국 미네소타 대학의 Schmidt-Dannert 그룹은 유사한 방법을 사용하였으나, PAL을 Rhodobacter shpaeroides의 tyrosine ammonia lyase (TAL)로, 나머지 유전자들은 Arabidopsis thaliana 유래의 유전자들로 대체함으로써 향상된 생산성을 보고하였다 [8]. 뉴욕주립대학의 Koffas 그룹은 효모인 Saccharomyces cerevisiae을 이종숙주로 사용함으로써 생산성을 향상시킬 수 있었다고 보고하였으나 [9], 이들 연구들은 재조합 미생물을 이용하여 flavonoid 화합물의 생산 가능성을 입증하였으나, 산업화를 위해서는 생산성이 낮으며, 보다 다양한 구조의 화합물의 생산은 성공하지 못하였으며, 또한 E. coli의 경우는 chalcone의 생합성 전구체인 malonyl-CoA의 세포내 농도가 낮아 생산성이 떨어지는 근본적인 단점이 있고, S. cerevisiae의 경우는 여러 개의 생합성유전자들을 발현시키는데 유용한 다중발현벡터 (multi-vector)의 구현이 어려운 단점이 있다.
본 연구팀은 S. coelicolor의 coenzyme A ligase (ScCCL)와 Arabidopsis thaliana의 chalcone synthase (atCHS)를 single ermE*를 사용하여 S. venezuelae에서 발현하여 naringenin과 pinocembrin의 생산을 확인하였다 [10]. 또한 이종숙주에 적합하도록 codon optimize하여 생합성한 flavone synthase I gene (Petroselium crispum)을 S. venezuelae에서 발현하고 flavanones을 넣어주었을 때, flavones이 생산되는 것을 확인하였다. 추가로 동일한 방법으로 optimize한 합성 유전자 3β-hydroxylase gene (Citrus siensis)과 flavonol synthase gene (Citrus unshius)을 S. venezuelae에서 발현하여 flavonols을 성공적으로 생산하였다 [11].
4. 토양미생물 및 해양미생물에서 방선균으로의 이종숙주 발현
토양미생물인 myxobacteria Sorangium cellulosum으로부터 생산되는 epothilne은 Taxol과 유사한 작용을 하지만, 부작용이 적어 내성이 있는 암세포에 강한 항암작용을 보여 차세대 항암제로 대두되고 있다. 그러나 자연생산균주인 Sorangium cellulosum은 성장속도가 매우 느리고 (doubling time, 16h) 생산성도 매우 낮으며, 유전자 조작이 어려워 유전자 조작에 의한 생산성 향상을 기대하기 어렵다. 이런 이유에서 많은 연구팀들이 이종숙주 생산시스템의 개발을 진행하고 있다. 미국의 Kosan Bioscience에 의해 E. coli에서 epothilone을 대상으로 시도되었으나, 수율이 1 μg/L 이하로 극히 낮았다 [12]. 또한 독일 Saarland 대학의 Rolf Müller group은 optimized synthetic gene을 이용하여 이종숙주 Myxococcus xanthus에서 epothilone을 발현하여 100 μg/L를 생산하였다 [13]. 방선균에서는 본 연구팀이 유전자의 효율적인 발현을 위해 다른 방선균과 비교하여 성장 속도가 2배 이상 빠르며, 유전자 조작이 용이하여 이종숙주로서 개발 가치가 매우 높은 S. venezuelae를 이종숙주로 선정하고 돌연변이주를 제작하여 거대 생합성 유전자 집단의 발현에 적합한 다중발현벡터 시스템 (multi-vector system)을 구축하였다. 이를 통하여 epothilone B 약 0.1 μg/L와 epothilone D 0.4 μg/L를 생산하였다 [14].
해양 천연화합물은 다양한 유용한 생리활성을 가지므로 경제적·산업적 가치가 매우 높으며, 현재 많은 수의 화합물들이 주로 항암제로서 전임상 또는 임상시험 중임. 그러나 보통 해양 생물의 조직으로부터 회수할 수 있는 양이 매우 제한적이므로, 임상시험에 필요한 충분한 화합물의 공급이 어려우며, 이는 해양 천연화합물로부터 신약개발의 가장 큰 문제점이다. 이의 원활한 공급을 위하여, 대량 생산하고자 하는 해양 천연물의 생합성 유전자집단을 해양 미생물로부터 확보하고 이를 유전자 조작 및 배양이 용이한 이종숙주에서 발현하는 이종숙주 생산시스템의 개발은 필수적이다. 해양 남조세균인 Moorea producens 유래 생리활성물질인 barbamide (약 28 kb)의 거대 생합성 유전자 집단을 이종숙주 생산시스템을 이용하여 생산성 향상 및 유도체 개발에 대한 연구를 진행하여, barbamide 거대 생합성 유전자 집단을 S. venezuelae에서 이종숙주 발현하여 4-O-demethylbarbamide의 생산을 확인하여 해양 천연물의 효율적 미생물 생산시스템의 개발 예로써, 완전한 marin cyanobacterial NRPS/PKS의 첫 이종숙주 발현이다 [15].
현재 본 연구팀은 S. venezuelae를 이종숙주로 선정하고 이종숙주에 codon optimize된 epothilone 생합성 유전자 집단과 해양미생물 유래 cryptophycin 생합성 유전자 집단을 발현, 생산성 향상을 위한 연구를 진행 중이다 (그림 3).
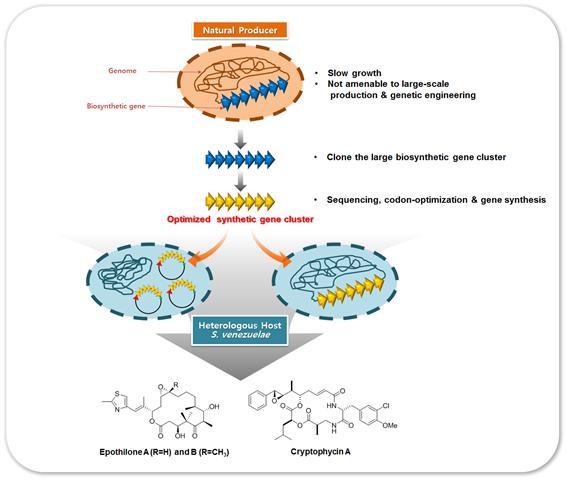
그림 3. S. venezuelae 이종숙주 생산시스템을 통한 Epothilone과 Cryptophycin의 생산
5. 참고문헌
[1] Wendt-Pienkowski E, Huang Y, Zhang J, Li B, Jiang H, Kwon H, Hutchinson CR, Shen B. Cloning, sequencing, analysis, and heterologous expression of the fredericamycin biosynthetic gene cluster from Streptomyces griseus. J Am Chem Soc. 2005 Nov 30;127(47):16442-52.
[2] Chen Y, Wendt-Pienkowski E, Shen B. Identification and utility of FdmR1 as a Streptomyces antibiotic regulatory protein activator for fredericamycin production in Streptomyces griseus ATCC 49344 and heterologous hosts. J Bacteriol. 2008 Aug;190(16):5587-96.
[3] Eustáquio AS, Gust B, Li SM, Pelzer S, Wohlleben W, Chater KF, Heide L. Production of 8'-halogenated and 8'-unsubstituted novobiocin derivatives in genetically engineered streptomyces coelicolor strains. Chem Biol. 2004 Nov;11(11):1561-72.
[4] Siebenberg S, Bapat PM, Lantz AE, Gust B, Heide L. Reducing the variability of antibiotic production in Streptomyces by cultivation in 24-square deepwell plates. J Biosci Bioeng. 2010 Mar;109(3):230-4.
[5] Jung WS, Lee SK, Hong JS, Park SR, Jeong SJ, Han AR, Sohng JK, Kim BG, Choi CY, Sherman DH, Yoon YJ. Heterologous expression of tylosin polyketide synthase and production of a hybrid bioactive macrolide in Streptomyces venezuelae. Appl Microbiol Biotechnol. 2006 Oct;72(4):763-9
[6] Park JW, Park SR, Nepal KK, Han AR, Ban YH, Yoo YJ, Kim EJ, Kim EM, Kim D, Sohng JK, Yoon YJ. Discovery of parallel pathways of kanamycin biosynthesis allows antibiotic manipulation. Nat Chem Biol. 2011 Oct 9;7(11):843-52.
[7] Hwang EI, Kaneko M, Ohnishi Y, Horinouchi S. Production of plant-specific flavanones by Escherichia coli containing an artificial gene cluster. Appl Environ Microbiol. 2003 May;69(5):2699-706.
[8] Watts KT, Lee PC, Schmidt-Dannert C. Exploring recombinant flavonoid biosynthesis in metabolically engineered Escherichia coli. Chembiochem. 2004 Apr 2;5(4):500-7.
[9] Yan Y, Kohli A, Koffas MA. Biosynthesis of natural flavanones in Saccharomyces cerevisiae. Appl Environ Microbiol. 2005 Sep;71(9):5610-3.
[10] Park SR, Yoon JA, Paik JH, Park JW, Jung WS, Ban YH, Kim EJ, Yoo YJ, Han AR, Yoon YJ. Engineering of plant-specific phenylpropanoids biosynthesis in Streptomyces venezuelae. J Biotechnol. 2009 May 20;141(3-4):181-8.
[11] Park SR, Paik JH, Ahn MS, Park JW, Yoon YJ. Biosynthesis of plant-specific flavones and flavonols in Streptomyces venezuelae. J Microbiol Biotechnol. 2010 Sep;20(9):1295-9.
[12] Mutka SC, Carney JR, Liu Y, Kennedy J. Heterologous production of epothilone C and D in Escherichia coli. Biochemistry. 2006 Jan 31;45(4):1321-30.
[13] Oßwald C, Zipf G, Schmidt G, Maier J, Bernauer HS, Müller R, Wenzel SC. Modular Construction of a Functional Artificial Epothilone Polyketide Pathway. ACS Synth Biol. 2012 Nov 5.
[14] Park SR, Park JW, Jung WS, Han AR, Ban YH, Kim EJ, Sohng JK, Sim SJ, Yoon YJ. Heterologous production of epothilones B and D in Streptomyces venezuelae. Appl Microbiol Biotechnol. 2008 Nov;81(1):109-17.
[15] Kim EJ, Lee JH, Choi H, Pereira AR, Ban YH, Yoo YJ, Kim E, Park JW, Sherman DH, Gerwick WH, Yoon YJ. Heterologous production of 4-O-demethylbarbamide, a marine cyanobacterial natural product. Org Lett. 2012 Dec 7;14(23):5824-7.









 방선균 이종숙주 생산시스템의 연구 동향
방선균 이종숙주 생산시스템의 연구 동향






