세포제어 인공 유전회로 연구동향
김근중 교수, 이진영 연구원
전남대학교
1. 개요
응용관점에서의 생물학 연구 분야는 기술이나 기법의 발달과 함께 진보되었다. 따라서 보다 정밀하고 경제적이며, 예측 가능하고, 목적에 따라 재설계의 여지가 큰 기술적인 진보는 생물학 발전에 있어서도 필수 불가결한 요소이다. 발효나 효소공정과 같은 전통적인 기술에서 벗어나 적극적인 방법으로 생체시스템에 기술을 적용시킨 과정은, 다루기 쉬운 이종 숙주에서의 유전자 조작 과정을 통해 원하는 산물을 과생산 하는 것에서부터 시작되었다. 이러한 과정의 결과물로 생체 구성요소로서 자연적인 대사물질은 물론, 비천연 화합물의 거의 대부분을 생체 시스템을 통해 생산할 수 있다는 사실이 확인되었다. 생리적인 현상이 지닌 전형적인 특성, 즉 원하는 시기에 원하는 산물은 적절한 수준으로 생산하며, 항상성을 유지하려는 경향은 이러한 목적에 큰 장애물임이 부각되기 시작된 것도 이러한 시기이다. 따라서 목적산물의 생산 경로나 조절에 중요한 기능을 담당하는 효소나 단백질을 암호화한 유전자의 조율(기능 관점에서의 변이 제작이나 발현양 조절)이 적극적으로 시도되었고, 결과에 따라선 생체 시스템이 지닌 자연대사 능력을 뛰어 넘는 우수한 결과들이 보고 되었다. 하지만 이러한 과정이 지닌 기술 적용 대상이 제한된 유전자나 단백질에 한정되고 이 외에 제어/통제되지 않는 다른 생체 시스템에 의한 부반응이나 간섭반응이 지닌 근본적인 한계로 인해, 전형적인 유전자 조작에 의한 산물 생산성은 원하는 수준을 담보하지 못하고 있다. 이러한 기술적인 한계를 극복하려는 연구 분야가 합성 생물학이다. 따라서 합성 생물학의 궁극적인 목표는 기존의 범위를 뛰어넘는 새로운 전략(세포 수준에서의 유전자 재설계나 합성)의 시도와 이를 접목한 수요자 중심의 맞춤형 세포(공장, 진단, 치료 세포 등)의 구현이다. 이러한 과정에서 환경/생리 인자(온도, 산소, 염, 양분, 개체인식, 세포통신)와 같은 외부요인이나 생리활성 물질(대사체, 전사체, 단백질, 에너지, 환원력)과 같은 내부요인의 탐지/인식/연산/제어 기능은 반드시 필요한 요소이다. 따라서 세포를 목적에 맞게 프로그래밍화하여 제어/조절할 수 있는 유전자 회로와 이를 통합/분석하는 논리 게이트(logic gate)의 개발은 필수적인 요소이다 (그림 1).
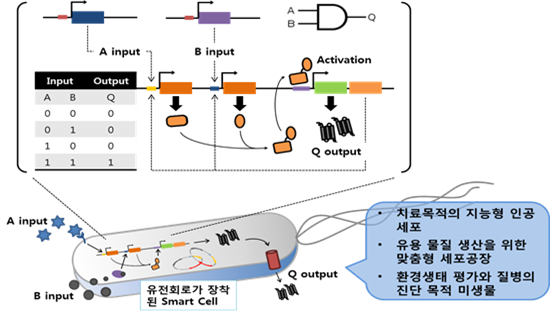
그림 1. 목적 지향성을 지닌 세포 제어용 유전회로와 이를 장착한 세포의 용도
이러한 과정의 주된 난제는 프로그래밍(세포의 성장, 대사, 분열, 사멸의 제어 및 통제)된 자동화 개념의 세포 제어 기술임으로, 유전회로 기술 구현은 기계/전기/화학분야와 달리, 규격화된 생산을 위한 공학적 체계의 수립이 어려워 발생한 생물공정의 높은 생산단가와 낮은 효율, 비균일한 제품의 질을 제고하는데 필수적일 수 밖에 없다. 따라서 센서, 스위치 및 구동 유전자를 조합하여 목표/논리체계를 갖춘 다중유전자 복합회로(하나의 입력 신호로 구동자가 거동하는 여러 개의 회로가 논리적으로 통합된 회로)를 구축하는 일은 전 세계적으로 매우 중요한 화두가 되고 있다.
2. 유전자 회로 제작 원리와 연구동향
유전회로 제작 연구는 생체 생리와 분자 생물, 생화학 현상의 통합적인 이해를 바탕으로 다양한 생체 소재(주로 생체 거대분자인 DNA, RNA, protein)에 공학적(예측, 설계, 표준, 규격화) 개념을 접목한 새로운 연구분야이다. 이러한 목적에 부합하는 산물로서 생체에 존재하는 새로운 소재를 발굴하거나 기 발굴된 발현이나 조절 시스템의 구성요소를 융합 혹은 재설계하는 방법으로, 매우 간단한 부품 수준의 모듈에서부터 복잡한 구조의 회로에 이르기까지 다양한 종류의 유전회로들이 개발되었다. 이때 사용되는 전형적인 회로 구성 요소는 유전자 전사나 단백질 발현 과정에서 흔히 이용되는 cis-acting element(promoter, operator, UP element, UTR, RBS, intervening and leader sequence 등)와 trans-acting element(regulatory proteins, peptide, micro RNA, ligand, interactive protein 등) 가 있다. 따라서 유전자 회로의 구성은 전자회로와 같은 입/출력 신호와 스위치, 논리/연산 인자, 회로화 등의 공학적 요소를 접목하지만 자연적인 생물생리가 지닌 복제나 전사, 번역과 같은 생명 현상의 중심원리(central dogma)에 따르는 조절원리를 이용할 수 밖에 없다. 따라서 현재까지 개발된 유전자 회로들은 전형적인 생체 조절 시스템이 지닌 입력 신호(물리화학적 자극 혹은 결합 인자)를 이용해 원하는 출력이 생성되는 지를 확인하는 patterning 이나 actuating 에 주된 목표를 두고 있다 (그림 2).
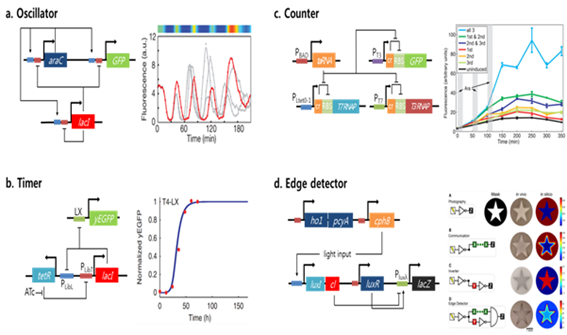
그림 2. 전형적인 유전회로의 종류와 설계구조에 따른 작동 원리 및 결과물
공학적 체계를 지닌 많은 부품들의 제작 과정이 그러하듯이 유전자 회로 제작과정에는 top-down 과 bottom-up 방식이 모두 활용된다. Top-down 방식은 생태계에 존재하는 생명체가 지닌 자연적인 유전자회로를 변형(치환, 축소 혹은 확장) 시켜 사용하는 것으로서 회로화된 시스템을 골격으로 원하는 패턴이나 구동에 필요한 요소들은 제거나 치환과정에서 기능성을 확인한 후, 재설계하는 방식이 주로 이용된다. 따라서 생태계에 기존재하는 회로화된 모든 시스템이 대상이 되며, 비교적 빠른 시간에 원하는 기능을 지닌 회로를 얻을 수 있는 장점이 있다. 이는 자연회로의 구성요소 모두의 기능을 알지 못하는 상황에서도 제거나 치환에 의해 달라지는 출력의 변화로서 원하는 패턴을 지닌 회로의 제작이 가능하기 때문이다. 반대로 bottom-up 방식은 회로 구성요소나 부품 개개의 알려진 특성을 바탕으로 원하는 패턴이나 거동이 가능한 회로 시스템으로 조립하는 방법을 이용한다. 따라서 상대적으로 적은 불확실성에 근거해 부분적으로 예측 가능한 논리를 회로로 접목할 수 있는 장점이 있으나, 구조나 기능이 예측 가능한 수준으로 충분히 알려진 소재의 수가 제한적이기 때문에 복잡한 논리를 구현하기에는 상대적으로 어려운 단점이 있다. 하기의 예에서와 같이 이러한 방법들로 다양한 유전회로와 관련시스템이 개발되었으나, 생체의 전형적인 회로를 주형으로 이용하는 방식이나 예측 가능한 부품을 이용하는 양쪽 모두의 방식에서 쉽게 해결할 수 없는 근본적인 문제점이 있다. 즉 알려진 자연회로나 특성이 규명된 부품, 소재의 대부분이 생체 시스템의 생리/생화학에 있어 중요한 기능을 담당하고 있기 때문에 다른 생체 요소와의 상호간섭이나 부반응, 자연적인 quality control (합성과 변형, 제거의 통제 기작)을 벗어나지 못하는 근본적인 단점이 남아 있기 때문이다.
세포제어나 조절 목적의 유전회로로는 toggle switch(Gardner et al., 2000)나 bi-stable circuit(Kim et al., 2006), oscillator(Stricker et al., 2008) 와 같이 비교적 단순한 개념을 구현한 예에서부터, 전기(전자)회로를 모티프로 timers, counters, clocks, logic processors, pattern detectors, intracellular communication module 과 같은 다양한 논리 연산이 가능한 분자 유전회로가 알려져 있다. 위와 같은 유전회로는 세포의 행동과 표현형의 제어를 위한 도구로 탑재되어 이용될 수 있는 것으로 알려져 있다. 따라서 유전회로 구축과 부품개발 연구 결과로써 다양한 전사시스템과 이의 조절 방법, 단백질이나 RNA 의 구조적 특성에 근거한 스위치 설계, 이를 세포 내 구동 패턴 구현으로 접목할 수 있는 연구 결과들이 존재하는 것을 의미한다. 이러한 과정의 대부분의 예는 상용화된 리포터와 패턴 논리 구현에 기존의 전형적인 회로요소(프로모터/전사인자/스위치)를 사용하는 것이다. 결과적으로 기저 수준 이상의 간섭이나 세포에 기존재하는 유사한 회로망과의 충돌이 근본적인 문제점으로 남아 있으며, 생체 분자의 고유한 수명(half-life)으로 인한 신호지연이나 중첩 등의 가능성은 상존함을 의미한다. 따라서 제작된 유전자회로가 숙주세포의 세포질 환경으로부터 독립되어 정확하고 재현성있게 기능이 유지되도록 하는 데는 아직 많은 어려움이 있다. 예를 들어 Hasty 등(Stricker et al., 2008)에 의해 개발된 oscillator를 살펴보면 이러한 문제점을 이해하는데 도움이 된다. Oscillator 회로 구성은 activator 와 repressor 두 단백질로 이루어져 있으며, 각각이 회로 유전자의 발현을 증가 또는 억제시키는 전사인자로 작동한다. 이때 각각의 유도체가 두 전사인자의 강도를 조절하는 변수로 이용된다. 각각의 activator, repressor, 리포터 단백질(형광 단백질)은 동일한 degradation tag을 달아 lifetime 을 줄여줌으로써 oscillation 의 해상도를 높였다. 그 결과 대장균에서 매우 정확하고 여러 세대까지 oscillation 이 유지되는 회로가 제작되었다(그림 2참조). 하지만 이 회로의 경우, 회로 구동을 위해 단백질이 발현되는데 많은 시간이 필요하며, 정확한 작동을 위해서는 최소 2개의 전사인자를 사용해야 하는 문제점이 있다. 또 다른 문제로 대장균 숙주의 구성요소와 의도되지 않은 상호작용이다. 예를 들면 두 전사인자는 동일한 degradation tag 을 달고 있지만, 단백질 각각의 folding rate(maturation time) 와 landscape, 전사체의 안정성이나 구조, post-translational modification에 의해 서로 다른 process 과정을 거치게 될 수 있으며, 그 결과 oscillation 되는 정도가 일정하게 유지 되지 않고 달라질 수 있다. 이러한 문제점은 모든 회로 구성요소가 숙주에 의해 인식/제어/간섭될 수 밖에 없는 부품으로 구성되었기 때문이다.
최근에 상기 문제의 해결책으로 이용될 수 있는 진일보된 비간섭 독립구동 가능 요소로 진핵세포 조절요소인 synthetic modular enhancer를 박테리아에서 구현(Amit et al., 2011)하였고, orthogonal genetic logic gate 의 구현을 위해 이종 세포 유래의 새로운 전자 조절 쌍을 대장균에 도입(Tamsir et al., 2011)하여 회로 고유의 기능에 독립성을 부여하는 연구 결과들이 발표되었다. 또한 새로운 스위치나 회로 조절요소로 활용 가능한 생체 대사물 인식 ribo-switch 들이 발견(항생제 인식 리보스위치, Jia et al., 2013; 에너지 인식 리보스위치, Watson and Fedor, 2012)됨으로써, 이를 기존 회로망에 직접 혹은 변형해 접목할 경우 보다 복잡한 논리나 조절 방법을 제공할 수 있는 가능성이 제시되었다. 그러나 자연 생태계의 생물이 지닌 생체회로는 매우 복잡하며, 네트웍(회로끼리 연결된)으로 구동하는 특성에 의해 완전한 독립성을 구현하는 것에는 한계를 지니고 있다. 따라서 현재까지 알려지거나 구현된 회로(망)을 이용해 생체에서 선택적으로 구동시켜, 원하는 시기에 필요한 논리를 구현하는 일은 매우 어려운 과제이다. 결국 자유자재로 통제가 가능하고 원하는 행태로 조절 가능한 비간섭/독립구동 가능 인공(재설계나 합성) 유전회로 구축이 절대적으로 필요하나, 실응용이 담보된 회로망의 구축은 아직 초보적인 시연 단계에 있다. 이는 제작된 전형적인 단위 회로나 다중 회로가 생체내의 복잡한 전자/번역 인자들을 공유하고 있는 문제점과 외부에서 공급되는 입력신호, 이를 해석하는 수용체들의 중복성에 따른 결과이다. 현재 제작된 회로의 경우, 전사 수준에서의 신호 지연도 큰 문제가 되는 것으로 알려져 있다. 이러한 문제들을 해결하는 일은 회로의 실응용을 위해 반드시 필요한 선행과제이다. 실응용의 궁극적 대상을 인체로 가정하고 질병진단과 치료를 목표로 하는 회로라면 다음과 같은 특성을 지녀야 하기 때문이다. 사람세포에서 질병탐침이나 진단, 이에 대응해 치료약물의 생산을 유도하는 유전회로의 경우, 입력 신호(질병마커나 특정 대사물의 농도 변화)에 고감도로 작동해야 하며, 정량성을 지녀야 하고, 생체 독성이 없어야 하고, 치료 후에 생체 독성을 지닐 수 있는 약물의 제어나 제거나 가능해야 하기 때문이다.
3. 실응용에 필요한 유전회로 구성요소
구동 패턴 분석에서 벗어나 실응용성을 갖춘 유전회로가 지녀야 할 특성으로, 신호지연의 문제점을 해결할 수 있는 민간성과 감도, 상호간섭이나 교란 등의 문제점을 피할 수 있는 독립구동, 전형적인 유전자 발현이 지닌 leaking(basal level expression)문제를 해결할 수 있는 on/off 화(전기 스위치와 같은 tight regulation 의미)가 매우 중요한 요소이다. 이러한 점에서 진핵세포와 유사하게 원핵 세포질도 고도로 구획화(localization and also sequestration)되어 있으며, 상호작용하는 인자들에 의해 RNA 나 단백질 수준에서의 집적화가 가능하다는 사실은 중요한 해결책을 제시하고 있다. 이는 회로 구성요소를 localization 혹은 interaction tag 에 의해 다른 생체 요소들과의 상호간섭을 피해 구획화가 가능하다는 것을 의미하기 때문이다. 또한 재조합 유전자의 RNA 나 단백질을 QC 를 담당하는 변형/분해효소를 피해 특정부위로 인도하여 안정성을 도모하거나, 근접 발현하여 기능 복합체의 효율적 회합(association)을 유도할 수 있다는 것을 보여준다. 이는 단백질 암호화 서열과 무관하게 회로 구성 요소의 세포 내 위치와 발현 양을 조절할 수 있다는 가능성을 제시하는 것으로서, 신호의 강건성이나 재현성 담보에 매우 중요한 고려 인자인 것이다.
실응용 회로가 지녀야 할 비간섭성과 독립구동은 orthogonal function 을 지닌 새로운 부품의 발굴로 해결할 수 있을 것으로 기대된다. 회로가 장착되는 전형적인 숙주와 다른 생리를 지닌 이종세포에서 발견된 새로운 발현 혹은 조절 논리를 지닌 요소를 회로에 도입함으로써 숙주의 발현/제어 시스템과의 상호간섭을 피할 수 있기 때문이다. 간단한 예로 회로 장착 숙주가 원핵세포인 경우, 진핵의 발현/조절 시스템을 도입하면 원핵 요소와의 상호충돌이나 간섭이 없어 외부 신호에 민감하게 반응하며, 생리적인 burden 이 상대적으로 적을 수 있기 때문이다. 진핵의 kinase 의존성 signal cascade 나 전사 조절인자(TF)의 도입은 이미 진행중인 것으로 알려져 있다. 또한 메타지놈으로부터 발굴한 새로운 자연회로들이 지닌 유일성(uniqueness)을 이용해 회로화 하는 작업을 진행중인 예가 알려져 있다.
실응용, 특히 인체에 적용하는 회로가 갖추어야 할 중요한 전제요건은 tight regulation 일 것이다. 이는 leaking 에 의한 생체독성의 문제나 원하지 않은 반응을 최소화 하는데 절대적인 요소이기 때문이다. 하지만 유전회로의 주형이 되는 생체회로의 대부분은 생리적인 필요(scout 기능 이나 빠른 적응을 위한)에 의해 일정 수준이상이 입력신호가 없는 상황에서도 발현하는 것으로 알려져 있다. 따라서 생체 회로들은 on/off 가 아닌 up/down 조절 방식으로 조절된다. 이러한 근본적인 문제의 해결책은 비자연 요소를 접목한 인공 유전회로의 완전한 합성일 것이다. 또한 입력신호에 대응하는 회로의 반응을 다단계의 check point 를 거쳐 작동하게 함으로써 해결할 수 있을 것으로 기대된다. 하기의 실례에서 생(인)체 적용을 목표로 실응용 회로 요소들이 도입된 연구 결과를 보면 상기의 문제점과 이에 따른 해결 노력이 분명하게 들어날 것이다.
첫 번째 예로 항생제 내성을 지닌 균주와의 공생 결과로 biofilm에 둘러싸여 항생제가 잘 듣지 않는 감염성 박테리아의 제어방법이다 (Lu and Collins, 2007; Kohanski et al., 2007). 회로화된 논리의 접목을 통해 기존의 고농도 항생제 또는 복합 치료제를 투여하는 대신 분자공학적으로 변형된 박테리오파지를 이용하는 방법을 개발하였다. 변형된 박테리오파지는 효과적으로 biofilm 을 분해하여 그 안의 박테리아를 죽일 수 있으며, 또한 항생제의 효과도 높일 수 있다. 그 과정을 살펴보면 다음과 같다. 용균성 박테리오파지에 DspB 라는 biofilm 을 분해할 수 있는 효소 유전자를 탑재하고 biofilm 내의 특정 세균에 감염시키면, 대상이 되는 박테리아가 용균되어 DspB 효소가 biofilm 내에 노출되게 된다 (그림 3A). DspB 효소는 근처의 biofilm matrix를 분해하고 내부의 박테리아를 면역세포 또는 치료제(항생제) 등에 노출시킨다. 또한 항생제 내성이 있는 균주에 대해서는 SOS response 를 방해하는 유전자를 파지 바이러스에 탑재하여 감염시켜, 항생제에 대한 민감도를 높여주어 쉽게 제어할 수 있다. 박테리오 파지는 종에 따라 특정 숙주에 대해서만 특징적으로 감염되는 특징을 갖기 때문에 무해한 human microbiome 에는 큰 영향을 미치지 않을 것으로 판단되며, 따라서 효과적인 치료 수단이 될 수 있다. 하지만 분자공학적으로 변형되어 처치회로가 장착된 박테리오파지가 인체에 어떤 영향을 미칠지 명확하지 않아 치료제로서 실제로 적용하기에는 보다 많은 연구가 필요하다.
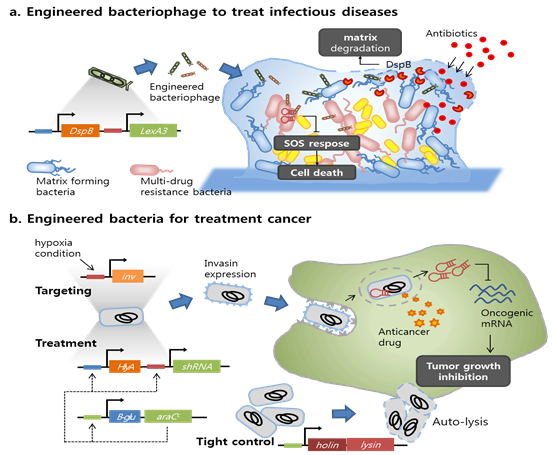
그림 3. 박테리아 감염과 암 치료에 실 적용하기 위한 유전회로의 예 및 구성요소
두 번째의 예로는 실응용을 전제로 개발한 유전회로를 장착한 세포를 이용하는 암세포 제어 방법이다. 지금까지의 암 치료 방법은 암세포뿐 만 아니라 정상세포까지 손상시키는 부작용을 지니고 있다. 또한, 기력이 약하거나, 너무 광범위하게 암이 퍼져 있는 환자의 경우 치료의 시도 자체가 불가능했다. 따라서 과학자들은 정상적인 세포와 암세포를 구별하여 암세포만을 특징적으로 공격하고 제어할 수 있는 방법을 찾기 위해 노력해 왔다. 그 결과 다양한 종류의 암세포에서 특이적으로 발현되는 유전자 또는 단백질 바이오마커를 확인하였으며, 암세포의 발달에 중요한 역할을 하는 oncogenic 유전자에 대한 지식도 축적되었다. 이러한 지식를 바탕으로 유전회로를 이용해 스스로 암세포를 찾아 공격하도록 프로그램된 대장균을 개발하였다(그림 3B). 개발된 대장균은 암세포 주위 환경과 같이 산소가 충분하지 않은 조건에서 invasin (inv) 이라는 Yersinia preudotuberculosis 유래의 접착성 단백질을 발현하여 endocytosis 를 통해 암세포 안으로 들어간다 (Anderson et al., 2006; Xiang et al., 2006). 이후 암세포 내에서 shRNA를 발현시켜 oncogenic 유전자의 발현을 차단해 암세포의 증식을 막는다. 하지만 환자의 혈관 속에서 혈압을 견디면서 면역 세포를 피해 움직이며 암세포에 도달해야 하고, 빠르게 invasion 을 발현해 암세포 내로 들어가야 하기 때문에 실제 이용을 위해서는 더 많은 개량이 필요하다. 비슷한 연구로 최근에는 박테리아로부터 유래한 minicell 에 bispecific antibody로 암세포 targeting 능력을 부여하고, 약물과 sh/siRNA를 탑재하여 연구한 예가 보고되었다(MacDiarmid and Brahmbhatt, 2011). 그 결과 항암제를 단독으로 처리할 때 보다 높은 약물 전달 능력을 확인하였으며, in-vivo에서도 높은 수준의 암 억제 효과가 나타나는 것을 확인하였다. 따라서 minicell 의 모세포나 minicell 자체에 회로화된 암세포 탐침 능력과 타겟팅, 약물생산 능력을 회로화시켜 구동시킬 경우 프로그램된 인공 세포를 이용하여 암세포를 선택적으로 치료할 수 있는 기술이 개발될 수 있을 것으로 기대된다.
4. 고찰
합성 생물학, 특히 유전자 회로에 주된 관점을 지닌 최근 연구 동향이 Science (Ruder et al., 2011; Nandagopal and Elowitz, 2011)에 특집으로 실렸다. 기고된 논문이나 주장에서 합성 생물학 기반 기술로 가장 중요한 것이 유전자 회로임을 명시하며, 현 단계의 중요 화두로서 단위 유전자 회로들의 유기적 통합(integration)이 필요한 시기가 되었음을 주장하고 있다. 또한, 실응용을 전제로 고도화된 회로가 장착된 목적지향세포의 전형으로 의료용 미생물이 중요함을 보여주고 있다. 유전회로는 구동 분석 목적의 패터닝 연구를 지양하고 실응용을 전제로 연구하면 응용범위와 학술적 가치가 매우 큰 연구분야이다. 현재는, 잘 알려진 유전자원을 토대로 설계와 재조합 과정을 거쳐 개발된 유전회로들을 이용해 생물학적 설계 원리를 이해하는 단계의 연구가 주류를 이루고 있다. 생체에서 구동시켜야 하는 목적으로 인해 생명현상의 중심원리를 벗어 나기는 매우 어렵다. 하지만 자연회로를 모방하되 공학적 개념과 작동 원리에 충실한 완전한 인공회로를 구축함으로써 생물을 이해하고 조절하며, 응용하는데 있어 가장 중요한 도구가 될 것이다.
5. 참고문헌
Amit, R., Garcia, H. G., Phillips, R., and Fraser, S. E. (2011). Building Enhancers from the Ground Up: A Synthetic Biology Approach. Cell 146, 105-118.
Anderson, J. C., Clarke, E. J., Arkin, A. P., and Voigt, C. A. (2006). Environmentally controlled invasion of cancer cells by engineered bacteria. Journal of molecular biology 355, 619-627.
Gardner, T. S., Cantor, C. R., and Collins, J. J. (2000). Construction of a genetic toggle switch in Escherichia coli. Nature 403, 339-342.
Jia, X., Zhang, J., Sun, W. X., He, W. Z., Jiang, H. Y., Chen, D. R., and Murchie, A. I. H. (2013). Riboswitch Control of Aminoglycoside Antibiotic Resistance. Cell 152, 68-81.
Kim, J., White, K. S., and Winfree, E. (2006). Construction of an in vitro bistable circuit from synthetic transcriptional switches. Mol Syst Biol 2.
Kohanski, M. A., Dwyer, D. J., Hayete, B., Lawrence, C. A., and Collins, J. J. (2007). A common mechanism of cellular death induced by bactericidal antibiotics. Cell 130, 797-810.
Lu, T. K., and Collins, J. J. (2007). Dispersing biofilms with engineered enzymatic bacteriophage. Proceedings of the National Academy of Sciences of the United States of America 104, 11197-11202.
MacDiarmid, J. A., and Brahmbhatt, H. (2011). Minicells: Versatile vectors for targeted drug or si/shRNA cancer therapy. Current opinion in biotechnology 22, 909-916.
Nandagopal, N., and Elowitz, M. B. (2011). Synthetic Biology: Integrated Gene Circuits. Science 333, 1244-1248.
Ruder, W. C., Lu, T., and Collins, J. J. (2011). Synthetic Biology Moving into the Clinic. Science 333, 1248-1252.
Stricker, J., Cookson, S., Bennett, M. R., Mather, W. H., Tsimring, L. S., and Hasty, J. (2008). A fast, robust and tunable synthetic gene oscillator. Nature 456, 516-U539.
Tamsir, A., Tabor, J. J., and Voigt, C. A. (2011). Robust multicellular computing using genetically encoded NOR gates and chemical 'wires'. Nature 469, 212-215.
Watson, P. Y., and Fedor, M. J. (2012). The ydaO motif is an ATP-sensing riboswitch in Bacillus subtilis. Nature chemical biology 8, 963-965.
Xiang, S. L., Fruehauf, J., and Li, C. J. (2006). Short hairpin RNA-expressing bacteria elicit RNA interference in mammals. Nature biotechnology 24, 697-702.
※ 다운로드는 로그인 후 이용하시기 바랍니다.









 세포제어 인공 유전회로 연구동향(전남대 김근중 교수, 이진영 연구원)
세포제어 인공 유전회로 연구동향(전남대 김근중 교수, 이진영 연구원)






