세포막 제어 기술의 적용을 통한 지방산 생산
김동훈 선임연구원
한국에너지기술연구원
1. 개요
바이오디젤, 계면활성제, 용제, 윤활유 등의 원료인 지방산에 대한 수요는 증가하고 있으나, 원료물질의 공급부족과 기존 공정의 비경제성에 따라 새로운 제조법이 필요한 상황이다. 기존의 식물자원을 이용한 지방산 생산은 넓은 재배면적을 요구하고, 전처리, 추출 시 다량의 에너지를 소모하는 문제점을 안고 있다. 이에 따라 최근 미세조류를 이용한 지방산의 생산에 대한 연구가 많이 진행되고 있지만, 추출 비용이 과다하고, 추출 시 다량의 유기용매를 사용한다는 환경친화적이지 못한 점, 유전자 조작이 용이하지 못하다는 단점이 있다. 이에 대한 대안으로 대장균과 광합성세균을 이용한 지방산 생산에 대한 연구가 각광을 받고 있다.
미생물을 이용한 지방산 생산 기술의 상용화를 위해서는 합성생물학을 통하여 미생물을 개량하는 upstream 공정(지방산 합성/불포화도 증대, 지방산 체외 분비 촉진)과, 제조된 맞춤형 미생물을 고속 배양하고, 및 배양액내 분비된 지방산을 분리하는 downstream 공정으로 크게 구분이 된다. 현재까지는 주로 upstream공정, 특히 지방산 분해 효소 억제를 통한 지방산 합성 증대와 thioesterase 발현, S-layer, peptidoglycan layer 생성 억제 등 세포막을 제어하여 지방산의 체외 분비를 원활하게 하는 세포막제어 연구가 활발히 진행되고 있다 [1,2]
2. 지방산 합성/불포화도 증대
미생물과 식물은 지방산 생합성에 관여하는 효소들이 모두 독립적으로 존재하여 개별적인 반응을 유도함으로써 지방산의 de novo synthesis를 매개하는 type II fatty acid synthase system을 가지고 있으며, 이는 지방산 생합성의 모든 단계의 효소가 하나의 복합체로서 작용하는 동물의 type I fatty acid synthase system과 대비된다 [3]. 따라서 미생물을 이용하여 지방산 생합성에 관여하는 효소들의 발현체계를 재구성하고, 결과적으로 지방산의 생성 정도 및 그 조성의 변화를 유도할 수 있다. 지방산의 생합성 속도를 증진시키기 위해서는 특히 지방산 합성의 초기 과정을 담당하는 효소들의 발현을 증진시키는 전략이 요구되는데, acetyl-CoA로부터 malonyl-CoA를 합성하는 acetyl-CoA carboxylase (AccABCD), malonyl-CoA를 malnoyl-ACP로 전환시키는 malonyl-CoA:ACP transacylase (FabD), 그리고 지방산 elongation 반응의 최초 전구체 C4-ACP를 합성하는 ß-keto ACP synthase III (FabH)가 그 대상이 된다. 이는 이들 효소가 다른 모든 탄소 대사작용을 매개하는 중심 대사회로로부터 지방산 합성 대사회로로의 metabolic flux를 전환시키는 역할을 담당하기 때문이며, 이후 지방산의 elongation 반응은 모두 ACP가 결합된 상태로 진행되어 다른 대사회로들과 구분되게 된다. 아울러 지방산 합성 증진을 위해서 탄소성 저장 물질인 poly-ß-hydroxybutyrate (PHB)의 합성 대사회로를 차단하는 등 세포의 성장에 필수적이지 않은 경로를 차단함으로써 지방산 합성 효율을 극대화 할 수 있다 [4].
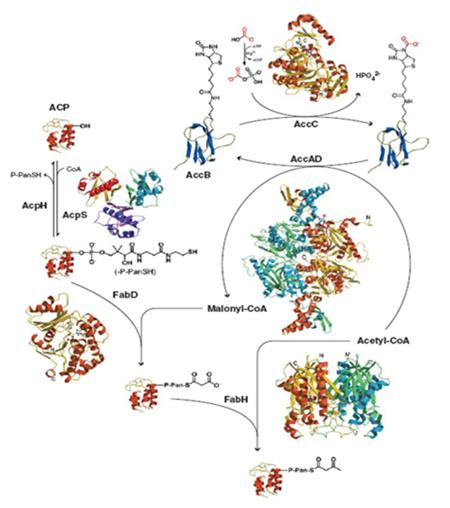
Fig. 1 지방산 초기 생합성 metabolic flow
지방산의 조성은 세포막의 물리적 성질에 해당하는 막의 유동성 (membrane fluidity) 및 막의 투과성 (membrane permeability)을 결정하는 주된 요인으로 작용하는데, 이러한 막의 물성이 변화하면 세포막에 존재하는 단백질의 기능에도 영향을 주어, 세포막의 주된 기능인 전자전달 (electron transfer), 세포 신호전달 (signal transduction), 물질의 선택적 투과성 등도 달라지게 된다. 지방산의 조성 중 세포막 환경에 가장 크게 영향을 미치는 요인은 지방산의 불포화도 (degree of unsaturation)이며, 지방산의 불포화화는 생체 내에서 서로 다른 두 종류의 방식으로 일어날 수 있다. 불포화 지방산은 최초 지방산 합성의 elongation 과정에서 ß-hydroxydecanoyl-ACP dehydratase/isomerase (FabA) 및 ß-keto ACP synthase I (FabB)의 연속된 반응을 통해서 합성되며 [5], 이미 합성된 포화지방산을 기질로 하여 fatty acid desaturase (desA/desB/desC/desD)의 작용을 통해서도 생성될 수 있다 [6]. 지방산을 산업적인 목적으로 생산하는 경우에도 지방산의 조성이 중요한 이슈가 되는데, 지방산의 생합성 속도뿐만 아니라 그 조성 또한 관련 효소들의 작용체계를 재설계하고 이를 제어함으로써 변화시킬 수 있다. 즉, ß-keto ACP synthase I (FabB)의 발현을 증진시키거나, 외래의 fatty acid desaturase를 도입함으로써 지방산의 불포화도를 높임과 동시에 부가가치가 높은 특정 불포화 지방산의 함량을 증가시키는 것도 가능하다.
3. 세포막 제어 기술
생리현상을 이해하는데 있어서 전자전달, ATP 합성, 광합성, 항상성 유지, 물질수송 및 신호전달 등의 주요한 기능을 담당하는 세포막에 대한 연구는 필수적이다. 그러나 기술적인 한계로 인하여 세포막에 대한 연구는 현재까지 그 결과가 미비하였다. 전체 단백질 중 약 40% 이상의 단백질이 세포막에 존재하는 막단백질이며, 이들 막단백질은 주변의 지질들과 상호작용하여 그 기능을 수행하게 된다. 세포막은 지질과 단백질 및 다당류 등으로 구성되지만 그 중에서도 막의 특성은 지질 및 지질을 구성하는 지방산의 조성에 따라 크게 영향을 받는다. 각종 오믹스 기술 및 생명정보학의 발전으로 마침내 인간에게 유용한 특정 기능을 효율적으로 나타낼 수 있는 맞춤형 미생물의 설계 및 제작이 가능한 시대에 이르렀다. 각종 유용 물질을 합성하도록 유전체를 재배치하고 유용 물질의 생산에 최적화된 생체를 구성하는데 있어 세포막을 제어하는 기술도 함께 요구되고 있다. 이는 세포막이 에너지의 생산, 물질수송 및 신호전달 등에 직접적으로 관여할 뿐만 아니라 다양한 효소들의 작용이 세포막 환경을 기반으로 이루어지고 있기 때문이다. 세포막을 구성하는 지방산 자체가 유용 물질로서, 우리는 지방산의 합성, 분해, 분비, 흡수 등의 대사과정 전반을 이해하고 이를 제어하여 특정 유용 지방산을 고효율로 생산하는 것을 일차적인 목표로 하고 있다. 뿐만 아니라, 세포막의 특성을 조절하는 기술개발을 통해 그 결과를 다양한 유용물질의 합성에도 적용할 수 있을 것이다
생성된 지방산은 체내에서 그 농도가 높아질 경우 새로운 지방산의 합성을 저해하는 feedback inhibition 현상을 일으키게 된다 [7]. 즉, 세포 내부에서 축적된 지방산은 지방산 합성과 분해와 관련된 전사조절인자인 FadR 및 FabR과 결합하여, 지방산 합성에 필요한 유전체의 발현을 억제하고 동시에 지방산의 분해 작용을 활성화시킨다. 따라서 이러한 feedback inhibition 현상을 억제하고 지방산 생합성의 효율을 높이기 위해서 생성된 지방산을 생체 외부로 분비 (secretion)시키는 전략을 고려할 수 있다. Thioestersae (TesA) 는 지방산과 ACP 사이의 에스테르 결합을 끊어주는 효소로서, 외래의 TesA를 세포 내에서 발현시킴으로써 합성된 지방산을 free fatty acids (FFA)로 전환하여 세포막 밖으로 분비되도록 유도할 수 있다 [8]. 이러한 전략을 채택할 경우 지방산 합성의 효율을 증진시킬 수 있을 뿐만 아니라 세포를 수득하고 유기용매를 사용하여 세포를 파쇄하지 않고도 배지 중으로 용출된 FFA를 수득할 수 있다는 장점이 있다. 아울러 long chain fatty acyl-CoA synthetase (fadD) 유전자를 파괴하여 이미 합성된 지방산을 분해하는 대사과정을 억제하고, 배지 중으로 용출된 FFA를 다시 흡수하는 작용을 담당하는 long chain fatty transporter인 FadL을 제거함으로써, 생산되어 세포 외부로 분비된 FFA가 세포로 다시 흡수되고 분해되어 이용되는 현상을 억제할 수 있어 세포의 지방산 합성 효율을 극대화 할 수 있을 것으로 기대한다.
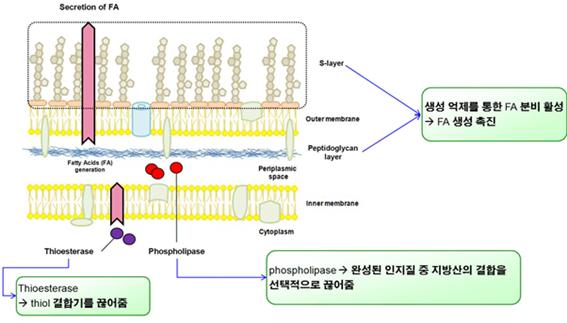
Fig. 2 세포막 제어를 통한 지방산 생산 증대 모식도
4. 참고문헌
(1) Lennen RM, Pfleger, BF. 2012. Engineering Escheria coli to synthesize free fatty acids. Trends Biotechnol. 30: 659-667.
(2) Liu X, Sheng J, Curtis R. 2011. Fatty acid production in genetically modified cyanobacteria. PNAS, 108: 6899-6904.
(3) White SW, Zheng J, Zhang YM, Rock. CO. 2005. The structural biology of type II fatty acid biosynthesis. Annu Rev Biochem. 74: 791-831.
(4) Lee IH, Park JY, Kho DH, Kim MS, Lee JK. 2002. Reductive effect of H2 uptake and poly-beta-hydroxybutyrate formation on nitrogenase-mediated H2 accumulation of Rhodobacter sphaeroides according to light intensity. Appl Microbiol Biotechnol. 60: 147-153.
(5) Feng Y, Cronan JE. 2009. Escherichia coli unsaturated fatty acid synthesis: complex transcription of the fabA gene and in vivo identification of the essential reaction catalyzed by FabB. J Biol Chem. 284: 29526-29535.
(6) Wada H, Murata N. 1990. Temperature-induced changes in the fatty acid composition of the cyanobacterium, Synechocystis PCC6803. Plant Physiol. 92: 1062-1069.
(7) Zhang YM, Rock CO. 2009. Transcriptional regulation in bacterial membrane lipid synthesis. J Lipid Res. Apr;50 Suppl: S115-119.
(8) Liu X, Fallon S, Sheng J, Curtiss R 3rd. 2011. CO2-limitation-inducible green recovery of fatty acids from cyanobacterial biomass. Proc Natl Acad Sci USA. 108: 6905-6908.









 세포막 제어 기술의 적용을 통한 지방산 생산(한국에너지기술원 김동훈 박사)
세포막 제어 기술의 적용을 통한 지방산 생산(한국에너지기술원 김동훈 박사)






