세포에너지 제어 바이오시스템의 개발 및 응용
김 필 교수
가톨릭대학교
1. 개요
세포 내에서 통용되는 에너지의 형태는 ATP 등을 포함하는 고에너지 인산결합, 막을 경계로 하는 이온농도구배의 일종인 proton motive force(PMF), 전자를 이동시키는 NAD(P)H 등이 존재한다. 통용되는 세포에너지는 외부의 빛 에너지나 혹은 유기물에 포함된 화학에너지의 대사과정으로부터 형성되어 세포 안에 축적된 이후 모든 생명현상에 사용된다. 에너지의 사용은 능동물질수송, 운동, 감각, 신호전달, 생장속도, 세포물질 생합성을 포함한다. 따라서 이러한 세포의 통용 에너지는 알려진 모든 생물시스템에서 공통적으로 사용되면서 생명현상에 사용되고 있으므로 모든 세포대사조절과 세포생리의 변화를 직•간접적으로 조절하는 인자이다 [1]. 세포에너지의 제어가 가능하다면 생명현상 전반에 걸친 제어가 가능할 것으로 추정되며, 최근 유전체 해독 기술과 유전체 수준에서의 유전자 조작이 가능해 짐에 따라, 인공적으로 설계된 생명체를 통한 고부가가치의 의약용 단백질이나 백신 등을 개발하거나, 연료, 플라스틱 등의 기존석유화학제품을 생물학적으로 합성하는 데 활용할 수 있다. 즉, 세포에너지를 인위적으로 활용하여 다양한 세포물질을 인공적으로 합성하는 과정을 촉진함으로써 기존에 기술적 임계점에 있었던 문제들(산화환원반응에 기반한 생물전환에서의 환원력 부족문제, 핵산활성형 당류소재의 부족문제, 세포 내 산물축적에 의한 product feedback inhibition 문제, 원료물질의 세포 내 수송속도의 제한문제, 외래단백질 발현의 경우 아미노산 부족이나 tRNA charging 속도의 제한문제)들을 돌파(breakthrough)할 수 있는 획기적인 선도기술을 확보할 수 있을 것으로 기대된다.
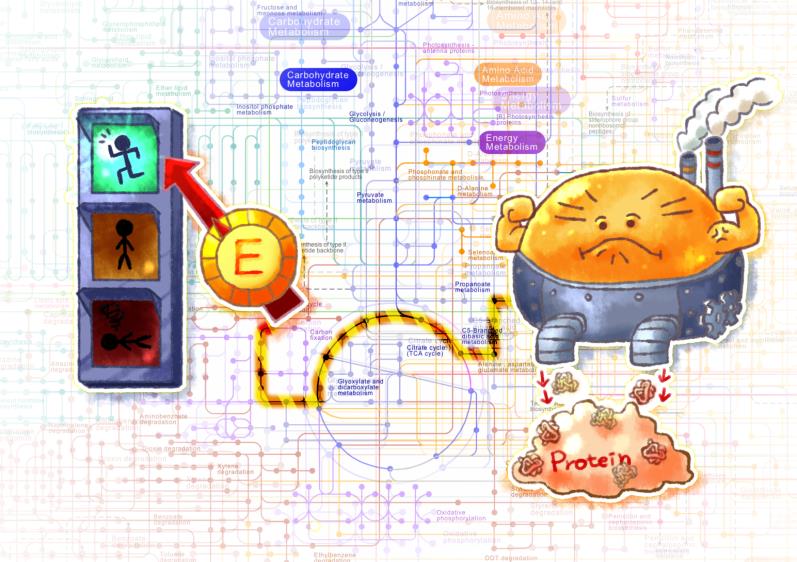
Figure 1. 세포에너지 제어를 통한 세포공장의 조절
2. 대사경로 변화를 통한 고에너지 세포모델해당과정(glycolysis)과 TCA회로를 연결하는 PEP carboxylase(Ppc)은 PEP의 고에너지결합을 야생형 대장균에서 열로써 방출하게 되는데, gluconeogenesis조건에서만 발현조절되는 PEP carboxykinase(Pck)를 인공적으로 발현하도록 조절하여 PEP로부터의 고에너지결합이 ADP에 전달됨으로써 ATP를 추가적으로 형성시키도록 인공적인 대사를 삽입하였다. 이때 대장균은 포도당과 CO2가 공급되는 조건에서는 야생형과 달리 세포 내 ATP 농도가 증가하였다 [2]. 또한 자발적인 Pck발현을 유도하고자 대장균의 게놈에서 Pck의 프로모터지역을 Ppc의 프로모터를 갖도록 대체함으로써 Ppc 발현되는 해당과정조건에서도 별도의 유도과정 없이 Pck가 스스로 발현되도록 조절하였다. 이렇게 인공적인 대사를 포함하는 균주는 야생형보다 ATP의 농도가 100% 증가된 것으로 관찰되었다. 따라서 플라스미드 또는 게놈조절을 통해 인위적인 Pck의 발현조절을 유도하여 열로 소실되는 PEP의 고에너지결합을 ATP로 저장함으로써 세포 내 ATP의 농도를 변환시킬 수 있는 대장균 모델을 구축하였다.
ATP의 인공적인 조절과는 별도로 세포의 환원력조절물질인 NAD(P)H의 농도를 변화시키기 위해게놈상에서 해당과정에 있는 phosphoglucose isomerase(Pgi)를 결손시킴으로써 탄소흐름을 오탄당인산회로로 유도하여 환원력이 야생형보다 높이 형성되도록 유도함과 더불어 낮은 수준으로 유지되는 NAD-dependent malic enzyme(MaeA)를 인공적으로 발현유도하여 TCA 중간체인 malate를 pyruvate로 변환시키면서 수반되는 NADH의 생성을 유도하였다. 이 결과로 세포 내 환원력 물질인 NADH와 NADPH의 농도는 야생형 대비 약 2배가량 증가되었고, 환원력 뿐 아니라 세포 내 ATP농도 역시 비례적으로 증가하기 되어 고환원력과 고ATP상태가 연동되고 있음을 확인하였음. 따라서 플라스미드 또는 게놈조절을 통해 대사경로를 변경함으로써 환원력분자인 NAD(P)H의 생성을 유도하여 고환원력 상태로 전환된 대장균 모델을 구축하였다 [3].
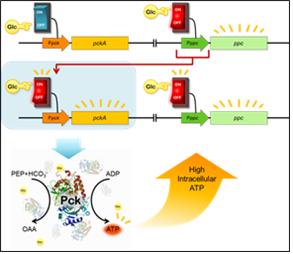
Figure 2. 인공적인 ATP 조절을 위한 전략
3. 고에너지 세포의 활용성
고에너지 대장균모델이 세포전구체의 생합성이 촉진되도록 조절되고, 단백질 합성기구인 리보솜의 농도가 높이 유지됨을 확인하였으므로, 산업용 단백질생산의 호스트로서 활용할 수 있다. 수용성인 형광단백질과 분비성인 Alkaline protease(AP)를 각각 야생형과 고에너지 대장균모델에서 발현하였을 때, 고에너지 대장균모델이 예상대로 높은 농도의 단백질을 생산함을 밝히고, 각각의 생장상태와 SDS-PAGE를 통한 단백질 농도를 검증하였다. 두 경우 모두 야생형 대비 2-3배가량 높은 수율로 외래단백질을 발현하는데 성공하였다. 외래단백질의 고발현은 세포의 생장이 고에너지에 의해 저해되는 시점에 최고치를 보였으며, 이후 세포생장이 회복됨에 따라 생산되었던 외래단백질이 점차 감소하는 경향을 보였다. 고에너지 상태인 대장균은 생장속도가 야생형보다 느리게 생장하다가 일정 시간의 낮은 생장속도 이후에는 생장이 회복되어 동일한 배지에서 최종적인 세포의 농도는 10-15%가량 증가하게 되었다. [4]
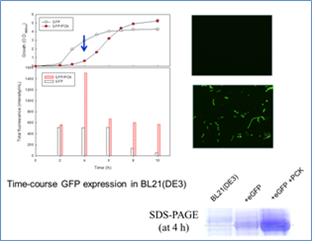
Figure 3. 고에너지세포에서의 단백질생산 증대효과
능동수송은 대사물을 농도구배에 역행하여 운반하는 수송기작이며, 세포에너지를 소비를 필요로 한다. 고에너지 대장균모델에서 능동수송에 의해 대사물 과생산에 의한 되먹임저해(feedback inhibition)을 극복할 수 있을 것으로 예측된다. 즉, 고에너지 대장균의 내부에 있는 에너지가 소모되어 외부로 대사물을 능동수송해 냄으로써 대사물의 생산공정과 고농도 대사물농도에 의한 스트레스 감소 등 산업적인 활용가능성이 있다는 사례를 제시한 바 있다.
이러한 활용가능성에도 불구하고, 세포에너지가 비정상적으로 높아지는 경우에 예측하지 못한 여러 가지 부작용으로 인한 산업적 활용성이 감소될 수도 있다. 즉, 세포의 에너지수준이 지나치게 높아지는 경우 세포의 생장속도가 억제되고 비정상적인 생리상태를 나타내므로 산업적 활용성을 제한할 수 있다. 제시된 문제를 극복하기 위해서는 보다 정밀한 수준에서의 세포에너지를 제어되도록 설계된 세포를 구축할 필요가 있다. 이를 위해 조절하고자 하는 개별 효소의 특성을 파악하여 [5] 그 발현제어를 위한 시스템을 개발하고 있다.
참고문헌
[1] Gaal et al., “Transcription regulation by initiating NTP concentration: rRNA synthesis in bacteria”, Science 278:2092–2097 (1997)
[2] Kwon et al., "A physiology study of Escherichia coli overexpressing phosphoenolpyruvate carboxylkinase", Biosci. Biotechnol. Biochem. 72: 1138-1141 (2008)
[3] Kim et al., "Alteration of reducing in an isogenic Phosphoglucose isomerase (pgi)-disrupted Escherichia coli expressing NAD(P)-dependent malic enzymes and NADP-dependent glyceraldehyde 3-Phosphate dehydrogenase", Letters in Applied Microbiology. 52(5): 433-440, (2011)
[4] Kim et al., "An engineered Escherichia coli having a high intracellular level of ATP and enhanced recombinant protein production" Appl Microbiol Biotechnol 94(4): 1079-1086 (2012)
[5] Lee et al., “Estimation of phosphoenolpyruvate carboxylation mediated by phosphoenolpyruvate carboxykinase (PCK) in engineered Escherichia coli having high ATP”, Enzyme Microb Technol. 53 (1): 13-17 (2013)









 세포에너지 제어 바이오시스템의 개발 및 용용(가톨릭대 김 필)
세포에너지 제어 바이오시스템의 개발 및 용용(가톨릭대 김 필)






