2개-인자 신호전달계를 이용한 인공신호전달계 개발동향
권오석
한국생명공학연구원
1. 개요
모든 생명체는 외부환경 변화를 신속 정확하게 인지하고 그에 따라 유전체 발현을 조절함으로써 끊임없이 변화하는 환경에 적응하여 생명을 유지하기 위한 다양한 신호전달기전 (signal transduction system)을 갖고 있다. 세균의 경우 2개-인자 신호전달계 (two-component signal transduction system)가 그 대표적인 예이며 대사, 생물막 형성, 병원성, 균밀도 조절을 포함한 거의 모든 세포 활성 조절에 관여하고 있다 (Hoch & Silhavy, 1995). 전형적인 형태의 2개-인자 신호전달계는 감지단백질 (sensor kinase)과 조절단백질 (response regulator)로 구성되는데 감지단백질은 transmembrane domain을 이용하여 세포막에 부착되어 있으며 외부 자극을 인지하는 감지도메인 (periplasmic sensing domain)과 신호를 받으면 ATP를 이용하여 특정 히스티딘 잔기를 자가 인산화하는 세포질 내 transmitter domain 으로 구성되어 있다. 조절단백질의 경우 감지 단백질에 의해 인산화되는 특정 아스파테이트 잔기를 포함하고 있는 receiver domain과 타겟 유전자의 프로모터에 부착하여 그 발현을 조절하는 helix-turn-helix DNA 부착도메인으로 구성되어 있다 (그림 1).
최근 미생물 유전체 정보 및 메타게놈 정보의 축적을 바탕으로 한 비교유전체 연구를 통해 미생물 신호전달계의 다양성과 광범위한 분포가 밝혀지고 있으며 대부분의 세균과 절반 이상의 고세균은 수십 종씩의 2개-인자 신호전달계를 갖고 있으며 평균적으로 세균 당 50여 종의 2개-인자 신호전달계가 존재하는 것으로 추산되고 있고 대표적인 모델 미생물인 대장균에는 30여 종의 2개-인자 신호전달계가 존재한다.
한편 2개-인자 신호전달계는 그 종류와 인지하는 신호가 다양하고, 각 시스템 간에 간섭 없이 신호 특이적으로 작동하며, 대부분의 경우 해당 숙주의 생존에 필수적이지 않으며, 조절하는 타겟의 수가 제한적이라는 특징을 갖고 있다. 아울러 2개-인자 신호전달계는 세균에서부터 식물에 이르기까지 광범위한 생물종에 분포하며 특정 숙주내로의 도입이나 제거가 용이하고, 구성요소들의 기능단위가 치환 가능한 모듈 성격을 갖고 있으며 나아가 최근 각종 단백질 관련 데이터베이스를 통해 신호전달계를 이루는 단백질의 도메인의 구조와, 분포, 단백질 구조, 및 도메인-도메인 상호작용에 관한 정보가 제공되고 있다. 따라서 2개-인자 신호전달계는 유전자 발현조절 및 네트워크 엔지니어링을 위한 좋은 소재로 간주되고 있다 (Ninfa, 2010). 본 보고서에서는 대표적인 예들을 중심으로 세균의 2개-인자 신호전달계의 합성생물학적 재설계에 의한 인공신호전달계 개발 연구동향에 대해 고찰해 보고자 한다.
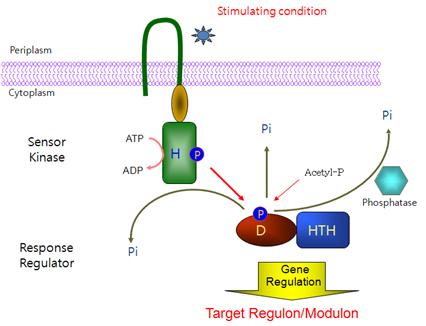
그림 1. 전형적인 형태의 2개-인자 신호전달계의 구성
인지단백질은 외부 신호를 인지하면 특정 히스티딘 잔기가 인산화되며 조절단백질은
인지단백질로부터 인산기를 받아 특정 아스파테이트 잔기가 인산화되어 전사 조절자로
활성화된다. Acetyl-phosphate와 같은 인산화합물에 의해 조절단백질이 인산화 되기도
하며 조절단백질의 인산기는 자극신호가 사라지면 다양한 방법에 의해 제거된다.
2. 신호전달계 단백질의 도메인 엔지니어링
대장균의 EnvZ-OmpR 2개-인자 신호전달계 대표적인 세균 유래 시스템으로써 각종 신호전달계 구성 단백질 도메인의 기능을 비교 연구하는 수단으로 가장 많이 활용되고 있으며 특히 세포막을 경계로 외부 환경 혹은 신호전달 물질 변화를 감지하는 감지도메인 연구에 많이 활용되고 있다(그림 2). 그 대표적인 예로 1989년 Inouye 연구팀에서는 대장균의 chemotaxis 시스템의 아스파테이트 수용체 Tar 가 삼투조절 EnvZ/OmpR 시스템의 EnvZ 감지단백질과 유사한 transmembrane topology를 갖는 사실에 착안하여 Tar 단백질의 리간드 인지 transmembrane 도메인에 EnvZ 단백질의 세포질 내 transmitter 도메인을 연결한 Tar-EnvZ 키메라 감지단백질 Taz를 제작하고 이 감지단백질이 아스파테이트를 인지하여 자가인산화되고 이어서 OmpR 조절단백질을 인산화시켜 타겟유전자 ompC의 발현을 촉진함을 입증하였다 (Utsumi, et al., 198). 이후 이 그룹에서는 리간드 부착에 따라 키메라의 자가인산화효소 활성과 OmpR-Pi 탈인산화 효소활성의 비율이 조절되며 신호감지도메인과 transmitter 도메인을 연결하는 linker도 키메라의 활성에 중요한 역할을 한다는 것을 입증하였다 (Yang & Inouye, 1993, Jin & Inouye, 1994).
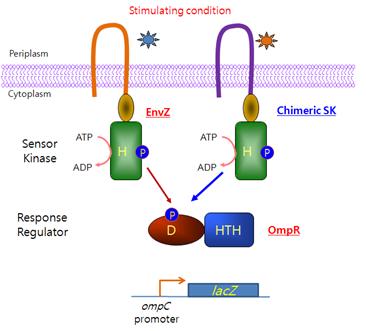
그림 2. EnvZ-OmpR 시스템을 이용한 인공신호전달계 개발체계
야생형의 EnvZ 인지단백질 (좌) 혹은 EnvZ의 transmitter 도메인을 포함하는
키메라 인지단백질 (우)과 OmpR 조절단백질간의 신호전달 및 타겟유전자 발현조절
또한 1994년에는 또 다른 chemoreceptor 인 Trg 단백질의 신호감지 및 세포막 함입 도메인과 EnvZ 단백질의 세포질 내 kinase/phosphatase 도메인을 조합한 Trg-EnvZ 하이브리드 단백질 Trz1을 제작되었으며 이 단백질의 신호감지 도메인이 리보즈가 결합된 리보즈결합 단백질을 인지하면 세포질 내 kinase/phosphatase 도메인의 활성을 촉진됨이 입증되었다(Bayumgartner et al., 1994). 이 결과는 신호감지 도메인에 의한 직접적인 저분자 리간드 인지 외에도 단백질과의 상호작용도 신호로 인지되어 융합 감지단백질의 활성을 조절할 수 있음을 입증한 예이며 나아가 신호전달계 골격이 아닌 제 3의 리간드 인지단백질의 엔지니어링에 의해서도 인공 신호전달계를 구축할 수 있는 가능성을 제시하였다.
한편 2002년 Ward 등은 nitrate을 감지하는 NarX 감지단백질의 periplasmic 감지도메인, 막함입 도메인을 Tar 케모리셉터의 세포질 내 신호전달 도메인과 연결한 NarX-Tar 하이브리드 단백질을 제작하여 이를 도입한 재조합 대장균이 야생형과 달리 nitrate나 nitrite에 주화성 반응을 보임을 입증함으로써 2개-인자 신호전달계의 감지단백질과 주화성 리셉터의 신호감지-전달 기전이 상호 호환될 수 있음을 다시 한 번 입증하였다 (Ward et al, 2002).
Periplasmic 감지 도메인을 갖는 일반적인 감지 단백질과 달리 세포질 내 PAS 도메인을 이용하여 세포막에 함입된 퀴논화합물의 산화/환원 상태를 인지하는 ArcB 인지 단백질의 세포막 함입 부위를 Tar 리셉터 단백질의 것과 치환한 Tar-ArcB 하이브리드 Tab 단백질을 제작하여 각종 신호 리간드에 따른 Tab-ArcA 신호전달 기전의 구동을 조사하였다 (Kwon et al., 2003). 그러나 다른 유사 하이브리드 인지단백질 들과 달리 Tab 하이브리드 인지단백질은 아스파테이트는 인지하지 못하는 반면 여전히 세포막에 함입된 퀴논화합물의 산화/환원 상태를 인지함이 확인되었다. 이 결과는 Tar와 ArcB는 상이한 신호인지 및 전달 기전을 갖고 있다는 것을 의미하며 이 후 유사 하이브리드 인지 단백질 제작의 한 기준으로 고려되고 있다.
이후 많은 2개-인자 신호전달계 감지단백질과 주화성 리셉터 단백질의 신호감지 도메인의 바로 뒤쪽 세포질 내 링커부위에 존재하는 HAMP (histidine kinase, adenylyl cyclase, methyl-accepting chemotaxis protein, phosphatase) 도메인이 외부 리간드 인지 도메인의 신호를 세포질 내 transmitter 도메인으로 전달하는데 중요한 역할을 함이 밝혀졌다 (Appleman et al, 2003). 또한 nitrate/nitrate 인지하는 NarX/L과 NarQ/P 시스템, 막스트레스를 인지하는 CpxA/R의 인지단백질들 간에 다양한 하이브리드 센서를 제작하여 그 구동을 조사함으로써 HAMP 도메인의 존재 여부 및 연결 부위가 이종 센서간의 하이브리드 센서 제작 시 고려해야할 주요 인자임이 확인되었다.
한편 동종의 세균에서 유래한 신호전달 기전뿐만 아니라 이종의 세균에서 유래한 인지단백질간의 하이브리드를 제작하고자 하는 시도로 2003년 Kumita 등은 토양세균 Sinorhizobium meliloti의 질소고정 관련 유전자의 발현을 뿌리혹 내 산소 분압에 따라 조절하는 FixL/J 시스템의 산소 인지도메인과 초고온균 Thermotoga maritima 의 ThkA 인지단백질의 transmitter 도메인에 연결한 수종의 키메라 CskA 단백질을 제작하여 산소분압 변화에 따른 자가인산화 변화를 조사하였다. 그러나 기대와 달리 CskA 단백질들은 산소분압에 따른 자가인산화 조절은 관찰할 수 없었으며 이는 인지단백질이 제 기능을 하기 위해서는 신호감지 도메인과 신호출력 도메인간에 정교한 상호작용 조절이 필요함을 시사함을 의미하였다 (Kumita et al, 2003)
대장균 NarX의 신호감지 도메인과 Myxococcus xanthus 의 집단활주운동 (social gliding motility)과 발달에 중요한 exopolysaccaharide 생합성을 조절하는 Dif 유사주화성 경로의 DifA 유사MCP 단백질의 신호 모듈을 연결한 NarX-DifA 키메라 NafA를 제작하여 M. xanthus 에 도입한 후 nitrate에 처리에 의해 Dif 경로가 유도됨을 확인함으로써 subdivision에 속한 세포 유래 신호감지-전달 단백질도 유사한 기전으로 운용됨을 확인하였다 (Xu et al, 2005). 이 그룹은 유사한 전략을 이용하여 NarX와 Frz 주화성 경로의 FrzCD 주화성 리셉터를 연결한 키메라를 이용한 연구결과도 보고하였다 (Xu et al, 2007).
2005년 Voigt 등은 EnvZ-OmpR 시스템에 광합성 세균 Synechocystis 의 파이토크롬 Cph1의 광수용체를 EnvZ 단백질의 세포질 내 transmitter 도메인에 연결한 Cph1-EnvZ 키메라 인공 광감지 단백질 Cph8을 제작하여 OmpR 조절 단백질과 연계하고 동시에 heme으로부터 광센서의 phycocyanobilin을 생합성하는 유전자 2종을 도입함으로써 적색광을 인지하여 타겟 유전자의 발현을 조절할 수 있는 대장균을 제작하였으며 (그림 3) 이 연구결과는 바이오부품을 이용한 합성생물학적 인공신호전달계 개발의 대표적인 예로 회자되고 있다 (Levkaya et al., 2005).
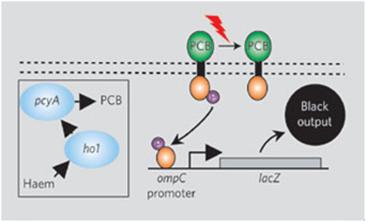
그림 3. Cph1-EnvZ 키메라 및 OmpR을 이용한 인공 광감지 시스템
(Levkaya et al., 2005)
한편 전술한 바와 같은 감지단백질 외에도 조절단백질을 엔지니어링을 통한 인공 신호전달계 개발 시도도 있었다. 전형적인 형태의 조절단백질은 쌍을 이루는 감지 단백질과 반응하여 인산기를 받아들이는 수용도메인 (receiver domain)과 신호에 따라 타겟 유전자의 발현을 직접 조절하는 DNA부착 도메인으로 구성되어 있다. 감지단백질에서의 예에서와 마찬가지로 조절단백질의 기능성 도메인을 치환하는 즉 한 조절단백질의 수용도메인과 다른 조절 단백질의 DNA 부착 도메인을 융합하는 방범이 주로 시도 되었다. 일례로 환경 내 무기 인 레벨을 감지하여 인포함 화합물의 유입에 관련된 Pho regulon 유전자의 발현을 조절하는 대장균의 PhoR/B 시스템의 PhoB 조절단백질과 아미노산 서열과 구조 측면에서 높은 상동성을 보이는 OmpR 조절 단백질의 도메인 융합을 통하여 키메라 조절단백질을 제작하려는 시도가 있었으며 수용 도메인과 DNA binding 도메인을 연결하는 링커 부위가 활성 융합단백질 제작에 중요함이 확인되었다 (Walthers et al, 2003). 한편 또 다른 Pho regulon 조절 시스템인 PhoR/P 시스템과 B. subtilis의 생존에 필수적인 YycG/F 시스템의 경우 PhoP의 수용 도메인과 YycF의 링커와 DNA binding 도메인을 연결한 PhoP'-'YycF 나 비슷한 전략으로 제작된 YycF'-'PhoP 하이브리드 조절단백질들의 경우 모두 활성을 가짐이 확인되었다 (Howell et al, 2003). 이런 결과들은 조절단백질의 체계적 조작을 통해 새로운 기능을 갖는 신호전달계를 제작할 수 있는 가능성을 제시하였다.
3. 인공 2개-인자 신호전달계 개발을 위한 새로운 시도
목적 지향적 신호전달계를 개발하기 위하여 인지단백질의 신호인지 도메인을 치환하는 방법 외에도 다양한 수준에서 기능성 모듈을 엔지이어링 한 예도 있는데 일례로 2003년 Hellinga 그룹에서는 단백질 구조 정보를 바탕으로 ribose binding protein의 리간드 인지 특이성을 재설계하여 높은 선택성과 친화도로 trinitrotoluene (TNT), L-lactate, serotonin을 감지할 수 있는 인공리셉터를 제작하였고 이를 전술한 Trz-OmpR 시스템과 연계하여 nM 수준의 TNT를 인지할 수 있는 인공신호전달계를 장착한 바이오센서를 개발하였다 (Looger et al., 2003). 흥미롭게도 이 시스템은 뒤에서 상세하게 설명할 식물체를 기반으로 한 합성생물학적 인공신호전달계 개발의 기본 원리로 사용되었다 (Antunes et al., 2011).
신호인지 도메인 전체를 치환하는 방법과 달리 특정 도메인이나 모티프를 도입하여 새로운 기능을 수행하는 신호전달계를 구축한 예도 있으며 Bradyrhizobioum japonicum의 FixL 감지 단백질의 산소 인지 heme-binding PAS (Per-Arnt-Sim) 도메인을 B. subtilis 의 YtvA 단백질의 LOV (light-oxygen-voltage) 광센서 도메인으로 치환한 융합단백질 YF을 제작하여 광 조사에 의해 융합단백질의 in vitro 인산화효소 활성이 제어되며 in vivo 조건에서도 광 의존적으로 타겟 유전자의 발현을 조절함을 확인한 경우가 해당된다 (Moglich et al., 2008). 나아가 이 연구에서는 LOV 도메인에 의해 감지되는 광신호가 링커부위를 40~60˚ 회전시켜 자가인산화 효소 도메인으로 전달됨을 확인하였다. 한편 최근 시카고 대학 연구팀이 보고한 희토류 금속 인지 시스템도 소규모 인지 모티프를 치환하여 신규 신호전달계를 제작한 예인데 이 경우에 사용된 살모넬라의 PmrA/B 시스템은 환경이나 숙주 내의 철이온을 인지하여 항생제 내성 관련 유전자 등 타겟 유전자의 발현을 조절하는 시스템으로 PmrB 감지 단백질의 30개 아미노산으로 구성된 periplasmic 감지 도메인에 iron binding motif를 갖고 있다. 이 연구팀은 PmrB 감지단백질의 철이온(Fe3+) 감지부위를 lanthanide-binding tag으로 치환하여 대장균에 도입함으로써 희토류계열 중금속의 일종인 란탄 (Tb3+)을 mM 수준에서 인지할 수 있는 인공신호전달계를 제작하는데 성공하였다 (Liang et al., 2013). 이는 PmrA/B와 같은 시스템을 기반으로 여타 금속이온의 생물정화나 흡착에 활용할 수 있는 합성생물학적 금속이온 감지시스템 구축 가능성을 입증한 예이다.
한편 콜로라도 주립대 연구팀은 대장균의 PhoR/B 시스템을 기본 골격으로 사용하여 인공 2개-인자 신호전달계를 개발하고 이를 식물체에 도입하여 TNT를 인지할 수 있는 phytosensor를 개발하였다 (그림 4). 이 인공 신호전달계의 구체적인 구성요소는 우선 전술한 TNT를 인지하도록 설계된 리보스 부착 단백질과, 이 단백질과 상호작용하는 Trg chemoreceptor의 신호감지 도메인에 PhoR 감지 단백질의 kinase 도메인을 연결한 Trg-PhoR 하이브리드 감지 단백질, PhoR과 쌍을 이루는 PhoB 조절단백질에 진핵세포 전사촉진 인자인 VP64가 연결된 PhoB-VP64 하이브리드 조절 단백질, PhoB-VP64 가 인지하여 부착할 수 있는 식물 세포용 프로모터와 여기에 연결된 표지 단백질 유전자 등을 포함하고 있으며 이 시스템을 이용하여 20 nM 수준의 TNT를 감지할 수 있음이 입증되었다 (Antunes et al, 2011). 이 연구결과는 세균에서부터 식물에 이르기까지 공통적으로 존재하는 2개-인자 신호전달계의 특징과 합성생물학적 재설계 가능성을 분명하게 보여주는 예라고 할 수 있다.
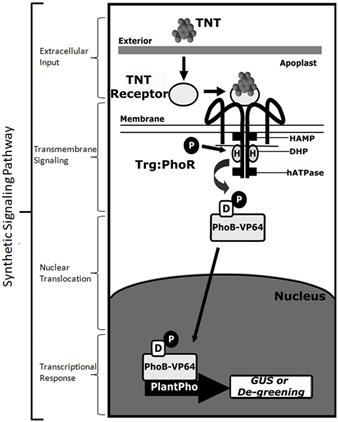
그림 4. 미생물 유래 2개-인자 신호전달계 골격을 이용하여 개발한 식물용 인공 신호전달계
(Antunes et al., 2011).
나아가 특정 도메인의 치환에 의존한 방법과 달리 고등생물의 스캐폴드를 이용한 신호체계에 착안하여 원핵세포의 신호전달 단백질에 진핵세포 유래의 단백질-단백질 상호작용 도메인을 연결하여 인산기 전이 방향을 재설정하려는 연구도 시도되었다 (Whitaker et al, 2012). 그 결과 스캐폴드를 이용한 in vivo co-localization을 통해 Taz 감지단백질 키메라와 원래 쌍을 이루는 조절단백질이 막 스트레스 감지 신호전달계의 CpxR 조절 단백질이나 구리이온 감지 신호전달계의 CusR 조절단백질 간의 인산기 전이 반응을 확인할 수 있었으며 인산화 도메인에 자가억제 기능을 부여하여 스캐폴드와 결합할 경우에만 인산기 전달 반응이 가능하도록 재설계 할 수 있음이 확인되었다. 이 결과는 비교적 잘 연구된 원핵 미생물 유래 신호전달계가 합성생물학적 설계의 좋은 타겟이 될 수 있음을 보여주는 예이다.
4. 고찰
2개-인자 신호전달계는 세균에서부터 식물에 이르는 다양한 생명체에 광범위하게 존재하고 적용될 수 있는 범용성, 한 세균 당 평균 50종 이상이 존재하는 다양성, 각 시스템이 감지하는 화학적/물리적 신호의 특이성, 구성 단백질이 기능성 도메인의 조합으로 이루어진 모듈적 특성, 그리고 쌍을 이루고 있는 인지-조절 단백질 간의 배타적이고 직교적인 관계 등으로 인하여 합성생물학적 인공신호전달계 개발 소재로 주목받고 있다. 그러나 지금까지 대부분의 2개-인자 신호전달계 재설계에 관련된 선행연구는 주로 특정 시스템을 기본 골격으로 기능성 도메인을 단순 치환하는 수준에 머물러 있었다. 하지만 최근 신호전달 단백질의 구조정보가 축적되고 생물정보학 기술이 발달함에 따라 점차 논리적인 설계와 co-localization 스캐폴드 활용과 같은 새로운 수단을 이용한 인공 신호전달계 개발 노력이 증대될 것으로 기대된다. 아울러 지금까지 주로 합성생물학적 개념의 검증 수준에서 수행되었던 신호전달계 개발도 점차 실용적 목적의 시스템 개발로 확대될 전망이다. 같은 맥락에서 특정 모델 미생물에 제한적으로 적용하던 수준을 벗어나 점차 산업용 미생물이나 효모나 식물과 같은 진핵세포, 나아가 최소유전체 세포나 2개-인자 신호전달 시스템이 존재하지 않는 동물세포 등에 적용할 수 있는 인공 신호전달계 개발이 급부상할 전망이다.
5. 참고문헌
1. Antunes MS, Morey KJ, Smith JJ, Albrecht KD, Bowen TA, Zdunek JK, Troupe JF, Cuneo MJ, Webb CT, Hellinga HW, Medford JI. Programmable ligand detection system in plants through a synthetic signal transduction pathway. PLoS One. 2011; 6:e16292.
2. Appleman AJ, Chen L-L, Stewart V. Probing conservation of HAMP linker structure and signal transduction mechanism through the analysis of hybrid sensor kinases. J Bacteriol. 2003; 185:4872–4882.
3. Baumgartner JW, Kim C, Brissette RE, Inouye M, Park C, Hazelbauer GL. Transmembrane signalling by a hybrid protein: communication from the domain of chemoreceptor Trg that recognizes sugar-binding proteins to the kinase/phosphatase domain of osmosensor EnvZ. J. Bacteriol. 1994; 176:1157–1163.
4. Hoch JA, Silhavy TJ. Two-component signal transduction. 1995. American Society for Microbiology, Washington, DC.
5. Howell A, Dubrac S, Andersen KK, Noone D, Fert J, Msadek T, Devine K. Genes controlled by the essential YycG/YycF two-component system of Bacillus subtilis revealed through a novel hybrid regulator approach. Mol Microbiol. 2003; 49:1639-1655.
6. Jin T, Inouye M. Transmembrane signaling. Mutational analysis of the cytoplasmic linker region of Taz1-1, a Tar-EnvZ chimeric receptor in Escherichia coli. J Mol Biol. 1994; 244:477-481.
7. Kumita H, Yamada S, Nakamura H, Shiro Y. Chimeric sensory histidine kinases containing O2 sensor domain of FixL and histidine kinase domain from a thermophile. Biochem Biophys Acta. 2003; 1646:136–144.
8. Kwon O, Georgellis D, Lin ECC. Rotational on-off switching of a hybrid membrane sensor kinase Tar-ArcB in Escherichia coli. J Biol Chem. 2003; 278:13192–13196.
9. Levskaya A, Chevalier AA, Tabor JJ, Simpson ZB, Lavery LA, Levy M, Davidson EA, Scouras A, Ellington AD, Marcotte EM, Voigt CA. Synthetic biology: Engineering Escherichia coli to see light. Nature 2005; 438:441-442.
10. Liang H, Deng X, Bosscher M, Ji Q, Jensen MP, He C. Engineering bacterial two-component system PmrA/PmrB to sense lanthanide iIons. J Am Chem Soc. 2013; 135:2037-2039.
11. Looger LL, Dwyer MA, Smith JJ, Hellinga HW. Computational design of receptor and sensor proteins with novel functions. Nature. 2003; 423:185-190.
12. Moglich A, Ayers RA, Moffat K. Design and signaling mechanism of light-regulated histidine kinases. J Miol Biol. 2009; 385:1433–1444.
13. Ninfa AJ. Use of two-component signal transduction systems in the construction of synthetic genetic networks. Curr Opin Microbiol. 2010; 13:240-245.
14. Utsumi R, Brissette RE, Rampersaud A, Forst SA, Oosawa K, Inouye M. Activation of bacterial porin gene expression by a chimeric signal transducer in response to aspartate. Science. 1989; 245:1246–1249
15. Walthers D, Ran VK, Kenney LJ. Interdomain linkers of homologous response regulators determine their mechanism of action. J Bacterioal. 2003; 185:317-324.
16. Ward SM, Delgado A, Gunsalus RP, Manson MD. A NarX-Tar chimera mediates repellent chemotaxis to nitrate and nitrite. Mol Microbiol. 2002; 44:709–719.
17. Whitaker WR, Davis SA, Arkin AP, Dueber JE. Engineering robust control of two-component system phosphotransfer using modular scaffolds. Proc Natl Acad Sci U S A. 2012; 109:18090-18095.
18. Xu Q, Black WP, Mauriello EMF, Zusman DR, Yang Z. Chemotaxis mediated by NarX-FrzCD chimeras and nonadapting repellent responses in Myxococcus xanthus. Mol Microbiol. 2007; 66:1370–1381.
19. Xu Q, Black WP, Ward SM, Yang Z. Nitrate-dependent activation of Dif signaling pathway of Myxococcus xanthus mediated by a NarX-DifA interspecies chimera. J Bacteriol. 2005; 187:6410–6418.
20. Yang Y, Inouye M. Requirement of both kinase and phosphatase activities of an E. coli receptor (Taz1) for ligand-dependent signal transduction. J Mol Biol. 1993; 231:335-342.
※ 다운로드는 로그인 후 이용하시기 바랍니다.









 2개-인자 신호전달계를 이용한 인공신호전달계 개발동향(생명연 권오석 박사)
2개-인자 신호전달계를 이용한 인공신호전달계 개발동향(생명연 권오석 박사)






