지능형 세포대사조절 시스템의 최근동향
: A dynamic sensor-regulator system
김선원 교수
경상대학교 응용생명과학부
서론
합성생물학은 인류가 원하는 미션을 부여했을 때에 이것을 받아서 스마트(smart)하게 처리하는 지능형 인공세포(Intelligent artificial cells)를 만드는 것이다. 즉 주어진 임무를 가장 최적의 방법을 이용해서 효율적으로 처리를 하는 세포가 지능형 인공세포라고 할 수 있다. 인공세포 중에서 부여된 임무가 소재물질의 대량생산인 경우에 세포는 하나의 화학공장으로 간주되고 세포공장(Cell Factory)이라고 불리게 된다. 결국 합성생물학은 식의약소재로 사용될 수 있는 고부가 기능성물질 또는 기존 석유유래 화학소재를 대체하는 기반화학물질을 최적의 효율로 대량생산하도록 세포의 대사경로를 지능형으로 재설계하는 것이다. 지능형 대사조절은 대사경로에 관여하는 유전자들의 발현이 대사환경 변화에 맞춰서 능동적으로 자율 조절되는 것을 필요로 한다. 이것은 기존 대사공학에서 유전자의 발현이 세포내/외부 대사환경 변화에 무관하게 프로모터의 세기, RBS(ribosomal binding site)의 세기, 발현벡터의 copy number에 의해서 결정이 되고, 한번 결정이 되면 세포내에서의 발현량이 고정되는 것과는 확연히 구분이 되는 것이다.
세포대사흐름의 다이내믹 콘드롤
상기의 지능형 대사조절의 개념은 University of California at Berkeley의 Jay D. Keasling 교수가 2010년 Cell 저널의 Leading Edge Essay 코너에서 제안을 했다. 그는 기존 발현벡터에서 유전자 발현패턴이 프로모터에 의해 일방적으로 결정되고 고정되는 것을 대사경로의 정적조절(Static control)이라고 하고, 세포내/외부 대사환경 변화에 따라서 조절되는 것을 역동적조절(Dynamic control)이라고 정의하였다. Keasling 교수는 대사경로가 dynamic control 될 때에 불필요한 중간대사물질의 축적 없이 대사자원이 효율적으로 목적산물 생산에 활용이 되고 낭비가 최소화 될 것이라고 예측했다. 그는 예로써 주변의 균체량을 인지하는 쿼롬센싱(Quorum sensing, 정족수인식)에 의한 대사경로 발현조절이 목적산물의 생산성 향상을 유도할 수 있다고 주장하였다(그림 1).
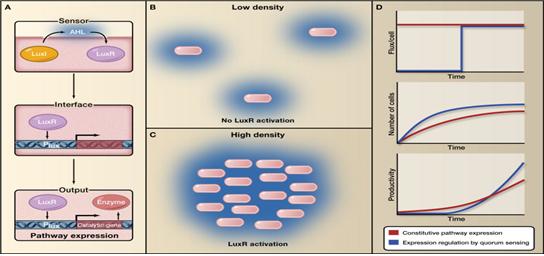
그림 1. 대사경로 다이내믹 콘트롤. 대사흐름 다이내믹 콘트롤은 대사경로 구성 효소들이 상시 발현되는 방식에 비해 더 높은 생산성을 얻을 수 있음. 상기 예의 다이내믹 시스템에서는 세포농도가 일정 농도에 도달할 때에만 대사경로 구성 효소들의 발현이 유도되는 것임. (A) 대사흐름 다이내믹 콘트롤러는 센서파트, 출력파트, 두 파트 사이의 연결파트로 구성됨. (B) 낮은 세포농도에서는 LuxI에서 만들어진 acyl-homoserine lactone의 농도가 낮아서 리셉터이자 프로모터 조절인자인 LuxR의 활성화가 안됨. (C) 일정 세포농도에 도달하면 LuxR이 활성화 되어서 Lux promoter (Plux)의 조절을 받는 대사경로의 유전자들의 발현이 유도됨. (D) 세포농도가 목적산물 생산에 적합한 농도에 도달했을 때에 생합성경로 유전자들의 발현이 더 높은 생산성을 만듬. (출처: Cell, Vol. 140, p19-23, Jan. 2010)
바이오센서를 이용한 유전자 발현조절
상기에서 제시된 다이내믹 콘트롤은 대사흐름을 능동적으로 조절을 할 수 있고 이것이 결국 생산성 향상과 연결이 될 수 있다는 것이지만 어떤 신호를 콘트롤의 지표로 삼을 것인가와 어떻게 선정된 신호를 인식(sensing)할 수 있는가 하는 것이 다음 문제로 제기된다. 세포가 일반적으로 인식할 수 있는 신호의 종류는 크게 3가지로 환경신호, 외부화학물질신호, 내부화학물질/대사물질신호로 이들을 인식하는 센서 시스템이 존재한다(그림 2).
환경신호센서(Biosensors for environmental signals): 세포는 다양한 환경신호들(빛, 온도, pH, 산소농도, 삼투압, 균체량)을 인식할 수 있고 이들을 인식하는 센서는 대부분 two-component system으로 되어 있다. Two-component system은 ① intracellular histidine kinase (HK) domain과 연결된 sensor domain으로 membrane-bound input domain, ② response regulator로 작용하는 intracellular output domain으로 구성된다. HK domains은 상동성을 갖고 있어서 이 domains과 이들의 cognate regulators를 스와핑(swapping)하면 환경신호에 반응해서 목표 유전자들의 발현을 조절하는 chimeric two-component systems의 제작이 가능하다.
외부화학물질센서(Biosensors for extracellular chemicals): 외부화학물질센서는 세포가 외부화학물질 자극에 반응할 수 있게 해준다. 대사공학에서 이들 센서는 배양 중에 외부화학물질 공급을 통해서 유전자들의 발현시기를 조절하는 것을 가능하게 한다. 아울러 목적산물 생합성에 2종 이상의 미생물이 필요한 경우, 즉 A 균주가 출발물질을 중간체로 전환해서 배출하면 B 균주가 받아들여서 최종산물로 전환하는 시스템에서는 중간체인 외부화학물질의 생산량을 인식해서 두 균주간의 대사활성을 조절하는 외부화학물질센서의 중요성이 더 높아진다.
내부화학물질/대사물질 센서(Biosensors for intracellular chemicals and metabolites): 세포내 화학물질, 특히 이종숙주에서 도입된 외래 대사경로에서 만들어지는 대사물질을 인식할 수 있는 바이오센서는 대사경로 조절에서 매우 중요하다. 이러한 센서는 대사경로 유전자들의 정교한 발현조절을 통해서 전체 대사과정이 균형을 이루고 대사자원이 효율적으로 이용되도록 하는 것을 가능하게 한다. 대표적인 예는regulatory RNA로 B. subtilis 등이 갖고 있는 riboflavin- or thiamin-responsive mRNA riboswitches를 들 수 있다.
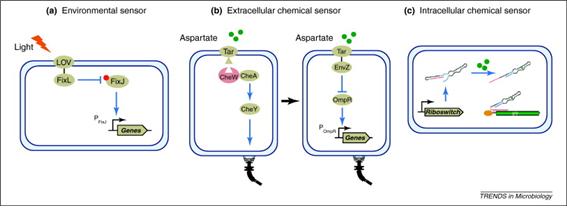
그림 2. Cellular biosensors 설계 및 제작. (a) Two-component system의 environmental sensor로 제작. Light-sensing domain (LOV from the B. subtilis protein YtvA)과 HK domain (FixL from the FixL-FixJ two-component system of B. japonicum)을 연결해서 blue light에 의한 유전자 발현조절. (b) Aspartate-sensing protein Tar의 intracellular domain을 EnvZ-OmpR two-component system의 EnvZ로 치환해서 대장균 chemotaxis signaling pathway로부터 extracellular aspartate를 인식하는 바이오센서를 제작. (c) Intracellular chemical theophylline을 인식하는 RNA-based biosensor 제작. Theophylline과 결합하고 구조적인 변화를 통해서 target mRNA에 결합해서 전사를 조절하는 riboswitch. (출처: Trends Microbiol., Vol. 19, p323-329, July 2011)
지방산 유래 바이오디젤 생산을 위한 dynamic sensor-regulator system
대사경로 최적화는 경로에 관여하는 유전자들의 발현 최적화와 일맥상통한다. 경로구성 유전자들의 발현이 너무 낮으면 생합성경로에 병목현상이 발생하고 반대로 너무 높으면 세포내 자원들이 불필요한 RNA, 단백질, 대사중간체들의 생산으로 낭비가 되어서 목적산물 생산에 이용될 수 있는 자원이 부족하게 된다. 더 나아가서 외래 단백질과 대사물질들은 숙주세포에 나쁜 영향을 미칠 수가 있기 때문에 이러한 독성 단백질들과 대사물질들의 세포내 축적은 생육저해 유발 또는 생산수율과 생산성의 저하로 귀결된다. 후자의 생산성 저하는 세포가 적대적인 환경에 적응하면서 발생하는 유전적인 변이가 주요 원인으로 저해현상을 유발하는 외래 대사경로 유전자들의 제거 또는 불활성화를 통해서 얻어진다. 상기 문제들을 해결하기 위한 promoters, intergenic regions, ribosome binding sites의 세기를 조절하는 몇 가지 유전자 발현조절 전략들이 개발되어왔다. 하지만 이들은 static control 방식으로 발현조절 시스템이 생물반응기 내의 특정조건에 맞추어져 최적화되어 있기 때문에 조건이 바뀌면 상기 유전자 발현조절 시스템은 적절하게 반응하지 못하고 최적의 생산성을 얻는 것은 불가능하다. 따라서 이상적인 유전자 발현조절 시스템은 세포 생리상태의 변화에 맞춰서 유전자 발현이 능동적으로 조절되는 것이다. 능동적인 유전자 발현조절(dynamic regulation)은 생명체가 세포내부 또는 외부환경 변화에 맞춰서 대사흐름을 조절하는 것을 가능하게 할 수 있다. 이것의 가장 바름직한 예는 핵심경로의 대사중간체 농도를 인지해서 능동적으로 대사중간체의 생산과 소모가 조절되는 지능형 대사회로를 구축하는 것이다. 이것은 세포 내/외부 환경변화에 맞춰서 대사중간체들이 적정 수준과 속도를 유지하면서 공급되게 함으로써 생합성경로가 항상 최적 생산성을 유지하도록 하는 것을 가능하게 한다. 상기의 지능형 대사경로를 구축하기 위해서는 생합성경로 상에서의 핵심중간체 생성량을 측정할 수 있는 센서와 최종산물 생산성이 최대가 되도록 경로를 구성하는 유전자 발현을 조절하는 대응조절자(cognate regulator)가 필요하다. 다행히도 생명체는 생합성 중간체 측정에 이용될 수 있는 다양한 세포내 물질들에 대한 센서를 진화과정에서 발전시켜왔다. 그러나 인공 생합성경로를 조절하기에 적합한 대응조절자는 자연계에 부족하다. 대부분의 조절자들이 인공경로가 아닌 목적산물과 무관하며 낮은 흐름속도를 갖는 자연경로에 맞춰서 진화되었기 때문이다. 따라서 인공경로를 최적화하고 목적산물의 생산성을 향상시키기 위한 DSRS (Dynamic Sensor-Regulated System) 구축은 좋은 인공조절자(dynamic regulator)를 설계제작해서 센서(natural sensor)와 연결하는 것이다.
UC Berkeley의 Keasling 교수 그룹은 상기의 DSRS 개념을 제시하고 대장균에서 지방산에 기반한 목적산물(바이오디젤) 생산에 성공적으로 적용하였다. 이 DSRS는 핵심 중간체를 인식하고 바이오디젤 생산에 관련한 유전자 발현을 역동적으로 조절하는 전사인자를 이용하고 있다(그림 3). 이 DSRS에 의해 바이오디젤 생산균주의 안정성이 증가했고 생산성은 DSRS 비적용 균주에 비해서 농도는 3배 증가한 1.5 g/l, 수율은 이론수율인 28%까지 향상되었다. 자연계에 다양하고 많은 바이오센서들이 존재하는 것을 고려하면 DSRS 전략은 다양한 생합성경로들에 적용되어 대사균형 최적화를 통한 최종산물의 생산농도와 전환수율을 증가 및 생산균주의 안전성 향상에 기여할수 있을 것이다.
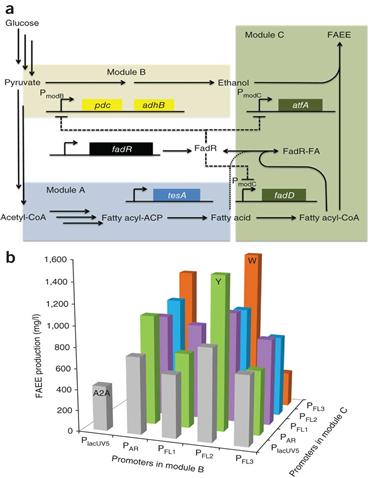
그림 3. (a) FAEE (fatty acid ethyl ester) 생합성경로를 3개의 모듈로 구성함. Module A는 대장균 유래 fatty acid synthase 유전자와 cytoplasmic thioesterase 유전자(tesA)로 구성되어 있고 지방산을 생산. Module B는 pyruvate decarboxylase 유전자(pdc)와 alcohol dehydrogenase 유전자(adhB)로 구성되어 있고 에탄올 생산. Module C는 acyl-CoA synthase 유전자(fadD)와 wax-ester synthase 유전자(atfA)로 구성되어 있고 최종산물인 바이오디젤 FAEE 생산. DSRS는 repressor 유전자(fadR)과 FadR-regulated promoters(PmodB와 PmodC)로 구성됨. 축적된 지방산이 없으면 FadR은 PmodB와 PmodC를 억제하고 에탄올과 acyl-CoA의 합성을 저해함. 반대로 지방산이 축적되면 chromosomal fadD로부터 발현된 단백질이 지방산을 fatty acyl-CoA로 전환하고, 이것은 FadR과 결합해서 FadR-FA로 되면서 FadR에 의해 억제되었던 에탄올과 acyl-CoA 생합성이 증가하고, 에탄올과 acyl-CoA를 중합해서 FAEE를 만드는 wax-ester synthase 발현도 증가함. (b) GC-FID에 의해 측정된 FAEE 생산량. FAEE 생산균주를 1 mM IPTG 첨가한 후에 37°C에서 3일간 배양. A2A는 DSRS가 적용되지 않은 지표균주. Module C에서 동일한 promoter를 갖는 균주는 동일한 색의 막대로 표시함. (출처: Nat Biotechnol., Vol. 30, p354-359, March 2012)
참고문헌
1. Holtz, W.J. & Keasling, J.D. Engineering static and dynamic control of synthetic pathways. Cell 140, 19-23 (2010).
2. Zhang, F. & Keasling, J.D. Biosensors and their applications in metabolic engineering. Trends Microbiol. 19, 323-329 (2011).
3. Zhang, F., Carothers, J.M., and Keasling, J,D. Design of a dynamic sensor-regulator system for production of chemicals and fuels derived from fatty acids. Nat Biotechnol. 30(4), 354-359 (2012).









 지능형 세포대사조절 시스템의 최근동향(경상대 김선원 교수)
지능형 세포대사조절 시스템의 최근동향(경상대 김선원 교수)






