고부가가치 단백질 소재를 암호화한 난발현 유전자 고효율 발현시스템 설계

전남대학교 김근중 교수
1. 개요
생물학분야의 태동이래 유전자발현에 관한 연구는 늘 연구중심에 서있는 중요한 화두이다. 기초생물학 분야에서부터 응용에 이르기까지 유전자 발현조절이나 기작, 논리의 이해는 세포를 제어하거나 조종하기 위한 바탕이 되기 때문이다. 따라서 유전자발현에 관한 새로운 기술이나 기법은, 세포의 근본적이며 내재적인 발현논리의 이해를 바탕으로 공학적 설계에 의한 발현품질 규격화나 표준화 (자연적으로는 난발현되나 재설계에 의해 일정 양 이상으로 인위적인 발현을 유도하는) 노력을 통해 다양한 연구그룹에서 시도되고 있다. 이러한 연구동향은 DNA, RNA, 그리고 단백질과 같은 생물소재를 표준화된 부품으로 이용할 수 있는 강건한 구동요소를 지니게 설계하고, 세포에 도입하여 목적산물의 생산이나 제어를 가능하게 하는 현 생물학의 핵심연구 분야 즉, 합성생물학의 주된 목표와도 잘 부합되는 주제이다 (Andrianantoandro, Basu et al. 2006). 이러한 합성생물학 연구를 통해 제작된 생물(주로 미생물)은 수 마이크로미터 내외의 공간(단일세포)에 도입된 체계적인 생물소재 설비를 이용하여, 기존의 석유화학을 이용하여 생산되던 각종 화학물질을 비롯한 소재/의약용 단백질 등, 다양한 목적산물 생산에 이용되기 때문에 세포공장 (cell factory) 이라 지칭하기도 한다. 이러한 과정의 성패여부는 전술한 유전자 발현의 규격화 요소를 지닌 표준화 작업이 결정한다.
일반적으로, 대부분의 생물산업에서 실응용 되는 생물유래 소재단백질 생산은 원균주를 직접 배양하여 정제하거나, 장기간의 복잡한 변이 유전자 선별과정 혹은 코돈 최적화에 따른 유전자 합성을 수반하며, 이러한 과정에는 특성이나 기작이 규명된 전형적인 발현인자를 사용한다. 하지만 잘 알려진 바와 같이, 이러한 노력은 전형적인 재조합 유전자 발현시스템이 지닌 한계 (세포내의 근본적인 유전자 발현논리를 이용하지 못하거나 공학적 요소를 지니지 못한 발현요소를 이용해서 생기는 문제)에 직면하게 된다. 특히, 일반적으로 이용되는 발현 모듈이나 회로는 세포 내 상호간섭이나, 기 존재하는 유사한 회로망과의 충돌이 근본적인 문제로 지적된다. 이러한 현상은 전형적인 유전자원을 이용하는 재조합 과정에 어떠한 요소들이 새로이 접목되어야 하는 지와 재설계의 바탕이 되는 생물논리의 이해가 무엇인지를 규명해야 하는 현 단계의 합성생물학 연구가 나아가야 할 지표를 제시한다. 본 기고문에서는 이러한 목적에 참고할 만한 새로운 유전자 발현논리에 기반한 발현모듈을, 난발현 소재 단백질을 암호화한 유전자 발현유도에 접목해 간단히 소개하고자 한다. 대상이 되는 다기능성 소재단백질은 최근에 새로운 생물소재로 각광받고 있는 하이드로포빈이다 (Erickson, Singh et al. 2011).
2. 다기능 단백질 소재 하이드로포빈의 산업적 가치
1991년 Schizophyllum commune에서 최초로 분리된 하이드로포빈은 아미노산 100~150개 정도의 작은 단백질로 주로 균사형 곰팡이에서만 발현된다. 진균류의 균사 형태형성과정에서 이러한 단백질은 소수성을 띄어, 단량체가 외부에 분비되면 자가조합 (self-assembly)이 일어나 소수성의 표층을 형성한다. 결과적으로 양친매성 다량체를 구성하여 균사 표면을 코팅한다. 이는 주변 환경변화에 대한 대응을 가능하게 함은 물론, 세포벽과 공기층 혹은 세포벽과 고체 표면사이의 계면에 위치하여 포자형성이나 자실체 발달, 그리고 감염구조 (숙주침입) 형성 과정에서 기능을 발휘하는 것으로 보고되어 있다 (Bayry, Aimanianda et al. 2012).
생물소재산업에서 일반적인 단백질에 비해 상대적으로 작은, 하이드로포빈에 주목하는 주된 이유가 상기한 특성에서 비롯된다. 즉, 하이드로포빈 고유의 소수성과 양친매성은 물체 표면에 소수성 코팅을 가능하게 하기 때문에, 안정되고 균일한 거품(기포, 기낭)이 필요한 화장품과 식품(맥주와 같은 거품 질감표현이 중요한 식음료 등) 제조에 적용할 수 있다. 또한 포유류를 비롯한 다양한 고등 생명체에서 면역회피 기능을 지니고 있기 때문에 의료용 코팅제로서의 개발 가능성도 있다. 더불어 강한 자기조합능을 지녀 식품의 민감한 생물학적 구성분을 포집해 생체적용 나노구조물 기능성 입자를 제조할 수 있어, 식품제조업이나 나노체 포장이 필요한 많은 산업분야에 차세대 혁신 기술로 대두되고 있다. 또한 화장품과 페인트 같은 나노기술의 응용에 의존하는 많은 영역까지 활용 가능성이 높아지고 있는 추세이다. 하지만 하이드로포빈 생산공정이 원균주인 곰팡이 자체를 이용하여 대량배양한 후, 정제하는 방법에 의존적인 것으로 알려져 있다. 이종숙주에서 하이드로포빈 생산이 어려운 이유는, 단백질 일차구조상에 존재하는 상보적인 8개의 시스테인 잔기가 3,4 차 구조를 형성함에 있어 필연적으로 정확한 이황화결합이 이루어져야 하고, 이들 단백질이 세포 내에 쌓였을 때 숙주세포 생장에 부담을 주는 이유에 기인하는 것으로 알려져 있다 (Linder, Szilvay et al. 2005).
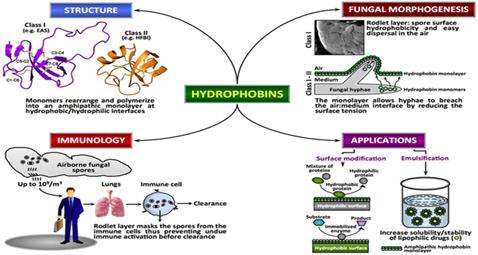
그림1. 하이드로포빈의 구조적 특성과 응용분야-출처:PLoS Pathg 8(5):e1002700
3. 난발현 재조합 단백질 발현 시스템 개선논리 및 설계
재조합 단백질 생산을 위한 발현 시스템들은 발현 벡터, 숙주, ORF 수준에서 접근이 시도되며, 관련되어 개발된 기술의 원형들은 비교적 잘 알려져 있다. 이러한 원형 기술들로 외래 유전자의 성공적 발현이 보고되고 있지만, 일반화 논리의 부족으로 인해 특정 유전자에 따라 재 최적화 하는 과정은 여전히 요구되고 있는 실정이다. 이를 해결하려는 노력으로 프로모터와 조절인자의 조합에 의한 전사체 양 증가 전략과 전사체의 2차구조(hairpin 이나 hammerhead 와 같은 2차구조로써 다양한 ribo-switch 도 포함된다), 열역학적 안정성 증가 위주의 발현 논리 개발이 이어졌지만, 여전히 모든 단백질에 적용 가능한 일반화 논리가 부족한 것으로 보고된다. 게다가 전사체와 전사체로부터 번역되는 단백질 양의 뚜렷한 상관관계가 없는 경우가 빈번하다는 문제는 아직도 해결해야 할 난제가 많음을 의미한다. 상기 문제의 해결책의 일환으로, 번역단계에서 발생하는 낮은 발현 효율은 숙주세포에서 잘 발현되는 고유유전자의 단백질 암호화 서열과 상이한 패턴을 보이는 코돈활용(codon usage) 빈도를 지닌 외래 유전자의 암호화 서열에서 기인한다는 분석 결과를 토대로, 코돈 최적화(codon optimization)라는 방법이 소개되어 일반적으로 적용되고 있다. 즉, 숙주세포 마다 서로 다른 코돈선호도를 통계적으로 분석하여 외래 유전자 내의 비선호코돈(rare codon) 비율을 감소시켜 번역속도를 증진시키는 방법으로 재조합 단백질의 발현량을 높이는 것이다. 그러나 야생형 ORF 염기서열을 동종의 아미노산을 암호화한 다른 서열로 바꾼 DNA로 인공적인 합성이 반드시 필요하므로 비용적으로 부담이 크고 암호화서열 분류 기준이나 방법에 따라 여러 종의 유전자를 합성해 실험해 보아야 하는 문제점이 있다. 게다가 기대와 달리, 야생형 유전자에 비해 향상된 발현량을 보이지 않는 경우가 빈번하게 관찰된다(Goltermann, Borch Jensen et al. 2011). 경우에 따라서는 극단적으로 codon de-optimization(최적화와 반대 논리를 적용해 선호 코돈을 비선호코돈으로 바꾸는 전략) 과정에서 발현량이 증가한다는 결과도 보고되고 있다. 코돈선호도 차이에 의한 재조합 단백질 발현효율의 우회적인 해결책으로, 해당 코돈(rare codon)에 대한 tRNA의 양을 보충하는 방법도 시도되었다 (Lee et al. .2006). 이 과정에서는 숙주세포에서 부족한 특정 tRNA를 코딩하고 있는 플라스미드를 숙주세포에 추가로 넣어주는 방법을 시도하였다. 그러나 상기 방법도 목적 유전자를 포함하는 벡터와 특정 tRNA를 코딩하는 플라스미드를 함께 제공하여야 하는 단점이 있다. 또한 dual vector-expression 의 근본적인 문제점인 세포 내 안정성과 이에 따른 재현성 있는 결과 도출의 어려움은 해결되어야 할 난제로 남아있다.
최근의 연구결과에 의하면, 단백질 발현과 관련된 새로운 논리(logic)들이 제안되고 있다. 우선 번역속도 조절이 중요한 변수가 되는 것 같다. 이는 RBS calculator 란 프로그램의 도입으로 설명되어지는 유전자 발현효율 증대로써 부분적으로 증명되고 있고, 희귀 코돈의 유전자 내 분포양상에 따라 많은 희귀코돈을 지닌 유전자의 발현효율이 예상과 달리 낮지 않다는 통계적 자료로도 설명되고 있다. 또한 단세포 미생물의 세포질도 구역화되는 양상을 보인다는 재미있는 현상도 단백질 발현에 참고해야 할 많은 시사점을 보여준다. 이를 기반으로 본 연구팀에서는, 전사체 번역에서 tRNA의 모집(recruiting)과 재사용(reuse)이 중요한 요소로 작용할 것이라는 가설을 세우고 비선호코돈을 재조합 단백질의 번역속도 조절용 태그(ramp tag)로 사용하는 방법을 고안하였다. Ramp 태그는 전사체가 번역과정에 들어가기 전에 속도를 조절하여 번역에 필요한 tRNA(비선호코돈이나 희귀코돈)를 충분히 불러들인 후에, 번역을 가속화하는 논리 태그이다. 또한 이러한 태그에 의해 불려 들인 tRNA 는 다음에 직면할 비선호(희귀)코돈의 번역속도가 줄어들지 않게 재사용됨으로써, 번역속도를 유지하는 기능을 지니고 있다. 이러한 태그는 코돈최적화 과정이나 변이 유전자 생성과정에 필요한 작업(유전자 합성이나 선별)이 필요하지 않다. 또한 유전자 본래 서열을 바꿀 필요가 없어 단백질 고유특성을 유지할 수 있을 뿐만 아니라, 인공합성이나 선별을 필요로 하지 않기 때문에 비용면에서도 경제적이다. 또한 태그라는 개념이 재조합이 필요한 외래 단백질의 유전서열에 따라 맞춤형으로 유동적인 적용이 가능하기 때문에, 적절한 과발현 태그 조합논리가 확립된다면 범용적(동/식물 세포주를 비롯하여 다양한 진/원핵 숙주에서의 단백질 생산)으로 활용이 가능할 것으로 기대된다. 선행연구를 통해 다양한 소재단백질이나 의약 단백질에 적용할 수 있는 과발현 유도논리와 태그를 확립하였고, 이를 하이드로포빈에 적용하는 연구를 진행중이다. 연구과정의 개략적인 설명은 아래 그림으로 대신한다 (그림 2).
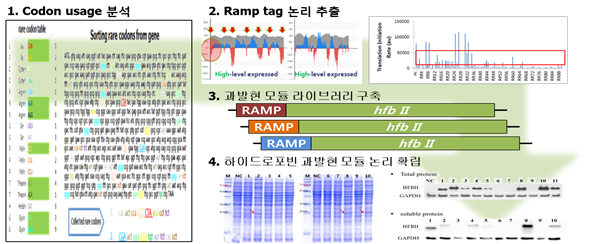
그림2. 하이드로포빈 과생산을 위한 과발현 태그논리 수립과 적용 예시
이와 더불어 하이드로포빈 과생산에 적용하고자 하는 새로운 발현논리로는 자가 발현 유도와 양성되먹임 조절원리를 이용한 단백질 발현 모듈이다. 하이드로포빈과 같은 산업화 단백질은 대량 배양 및 연속배양의 공정화가 필수적이기 때문에 세포 농도를 인식하여 목적 단백질의 발현시기를 자가 조절하는 quorum sensing(QS) 기작과 초기 자극(triggering) 후, 되먹임 조절에 의해 단백질 발현양을 증진(유지)시키는 양성되먹임 모듈이 적용되면 경제적이면서 효율적인 산업화 단백질 과생산 시스템이 확립될 것으로 예상된다. 새로운 발현논리를 지닌 발현 시스템을 장착한 모듈개발은 소재로서 단백질의 산업적 성공가능성 제고에 반드시 필요한 요소이다. 이는 자연적인 발현원리를 모방하되 공학적 개념과 작동 원리에 충실한 완전한 인공 발현모듈을 구축함으로써 구현할 수 있을 것이다. 이러한 발현모듈은 전형적인 바이오 시스템이 지닌 단점, 즉 여타의 연구분야보다 투자대비 낮은 효용성을 지니며 규격화된 생산을 위한 공학적 체계수립이 어려운 문제를 해결하는 방법이기도 하다. 따라서 세포농도 인식 자가발현 유도 모듈(Qurum sensing 논리이용)과 세포 기억현상의 공학적 유도가 가능하여 동기화에 의한 발현품질 유지에 매우 중요한 자가양성 피드백 모듈(유도체 비의존적 전사인자 이용)은 합성생물학에서 추구하는 기존의 전형적인 발현 시스템이 지닌 한계를 극복할 수 있는 새로운 발현 시스템으로 평가된다 (Hirsh and Schleif 1977, Guzman, Belin et al. 1995, Miller and Bassler 2001). 하이드로포빈 과생산에 이용될 새로운 발현논리를 지닌 발현모듈과 회로의 구성요소와 구동 기작은 아래 그림과 같다.
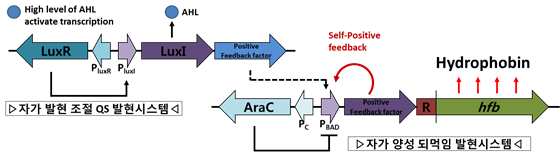
그림 3. 하이드로포빈 과발현을 위한 자가유도/양성 되먹임 발현모듈 모식도
4. 고찰
하이드로포빈은 양친매성 단백질로서 강한 자기조합능을 지니고 있고, 인체에 무해한 곰팡이로부터 유래되어 식품이나 의약품과 같은 생체 적합성이 필요한 분야는 물론, 나노크기의 기능성 입자로 도입될 수 있어 포장이나 약물 전달, 코팅, 발포제를 포함하여 많은 산업부분에 차세대 혁신 소재로 활용되고 있다. 또한, 화장품과 페인트 같은 나노기술의 응용에 의존하는 많은 영역까지 적용점이 증가하고 있는 추세이다. 현재의 생산 기술은 원균주인 곰팡이 자체를 이용하는 것으로 알려져 있어 재조합 미생물에서의 대량생산은 충분한 플랫폼 기술로서는 물론 제품으로써의 상업적 가치를 지닌다. 더불어 하이드로포빈 과생산에 이용된 과발현 모듈은 차세대 단백질의약품이나 친환경 화학소재/에너지를 생산하는 고효율 바이오융합 원천기술에 접목될 수 있다. 이들 요소를 활용한 세포공장구현은 생물학 전 분야에 걸쳐 발현유도 시스템의 활용을 극대화할 수 있는 기반을 제공할 수 있을 것이다.
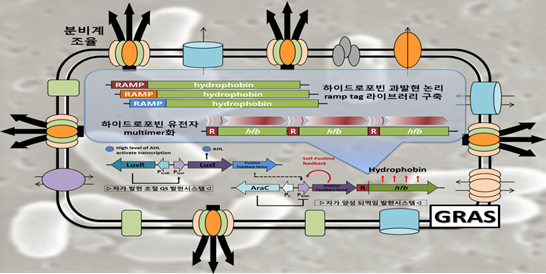
그림4. 난발현 단백질의 과발현 논리를 지닌 산업용 소재 단백질 과생산 세포 모식도
5. 참고문헌
1. Andrianantoandro, E., S. Basu, D. K. Karig and R. Weiss (2006). "Synthetic biology:
new engineering rules for an emerging discipline." Mol Syst Biol 2: 2006 0028.
2. Bayry, J., V. Aimanianda, J. I. Guijarro, M. Sunde and J. P. Latge (2012).
"Hydrophobins-unique fungal proteins." PLoS Pathog 8(5): e1002700.
3. Erickson, B., R. Singh and P. Winters (2011). "Synthetic biology: regulating industry uses of
new biotechnologies." Science 333(6047): 1254-1256.
4. Goltermann, L., M. Borch Jensen and T. Bentin (2011). "Tuning protein expression using
synonymous codon libraries targeted to the 5' mRNA coding region." Protein Eng Des
Sel 24(1-2): 123-129.
5. Lee SJ, Kim HS and Lee EY (2006). " Enhanced Heterologous Expression of Aspergillus
niger Epoxide Hydrolase and Its Application to Enantioselective Hydrolysis of Racemic
Epoxides" J. Korean Ind.Eng.Chem 12(5):557-560
6. Goodman, D. B., G. M. Church and S. Kosuri (2013). "Causes and effects of N-terminal
codon bias in bacterial genes." Science 342(6157): 475-479.
7. Guzman, L. M., D. Belin, M. J. Carson and J. Beckwith (1995). "Tight regulation, modulation,
and high-level expression by vectors containing the arabinose PBAD promoter." J Bacteriol
177(14): 4121-4130.
8. Hirsh, J. and R. Schleif (1977). "The araC promoter: transcription, mapping and interaction
with the araBAD promoter." Cell 11(3): 545-550.
9. Linder, M. B., G. R. Szilvay, T. Nakari-Setala and M. E. Penttila (2005). "Hydrophobins: the
protein-amphiphiles of filamentous fungi." FEMS Microbiol Rev 29(5): 877-896.
10. Miller, M. B. and B. L. Bassler (2001). "Quorum sensing in bacteria." Annu Rev Microbiol
55: 165-199.









 고부가가치 단백질 소재를 암호화한 난발현 유전자 고효율 발현시스템 설계
고부가가치 단백질 소재를 암호화한 난발현 유전자 고효율 발현시스템 설계






