고감도 이소프렌 감지 바이오센서의 개발 및 적용
한국생명공학연구원 김성근, 김하성, 이대희*, 이승구*
Kim SK, Kim SH, Subhadra B, Woo SG, Rha E, Kim SW, Kim H, Lee DH*, and Lee SG* (2018). A Genetically Encoded Biosensor for Monitoring Isoprene Production in Engineered Escherichia coli. ACS Synthetic Biology, 2018, 7(10):2379-2390 “Most-Read Article”로 선정 (2018.10.22.-2018.11.30)
1. 연구배경
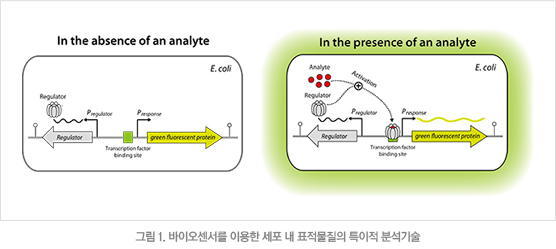
바이오매스를 원료로 이용하여 바이오에너지 및 유용 화학물질을 생산하는 연구는 미생물 생명공학의 오랜 관심사였다. 최근 대사공학 및 합성생물학과 더불어 유전자 합성기술이 비약적으로 발전함에 따라 다양한 미생물 유전체를 필요에 따라 설계하고 제작하는 것이 가능해지고 있다. 다양한 재조합 균주들의 생산성을 평가하기 위해서는 크로마토그래피와 같은 분석 장비들이 이용되는 데 이때 시간과 비용이 많이 드는 어려움이 있다. 이에 대한 해결책으로 전사 인자를 이용한 바이오센서가 주목을 받고 있다. 바이오센서는 분석 물질(analyte)에 결합하는 전사 인자(regulator)와 이 전사 인자에 반응하는 프로모터(Presponse) 및 리포터 유전자(fluorescent protein)로 구성되어 있다(그림 1). 즉, 분석 물질이 존재할 경우 전사 인자가 활성화되어 리포터 유전자인 형광 단백질의 전사를 개시하고, 이때 나오는 형광을 측정하면 단일 세포에서 수 μM로 낮은 농도의 물질도 빠르게 정량 가능하다. 본 연구에서는 합성 고무의 원료가 되는 이소프렌을 감지할 수 있는 바이오센서를 제작하고, 최적화하여 재조합 대장균에서 생산되는 이소프렌을 감지하고자 하였다.
2. 연구내용
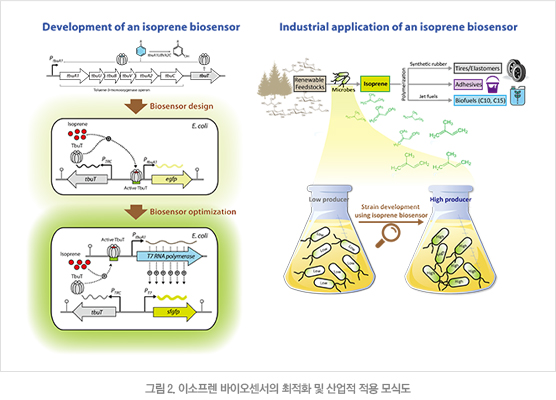
본 연구진은 이소프렌을 고감도로 감지하는 바이오센서를 개발하였고 이를 이용하여 대장균에서 생산되는 이소프렌을 쉽고 빠르게 정량하는 기술을 구축하였다(그림 2). 이를 위하여 먼저 이소프렌을 감지하는 전사 인자로 TbuT 단백질을 이용한 바이오센서 유전자 회로를 디자인하였다. 이어 바이오센서의 출력을 T7 RNA 중합효소(T7 RNA polymerase)에 의한 증폭반응으로 강화하여 이소프렌에 대한 민감도를 증가시켰다. 상기 방법에 의해 최적화된 바이오센서를 다양한 생산 균주에 적용한 결과 이소프렌 생산량과 비례하여 바이오센서의 신호가 5배 이상 증가함을 확인하였다. 이 이소프렌 바이오센서는 다수의 생산 미생물들로부터 고효율로 이소프렌을 생산하는 산업 미생물 및 합성효소의 선별에 이용될 예정이다.
2.1 이소프렌 바이오센서를 위한 유전자 회로 설계 및 제작
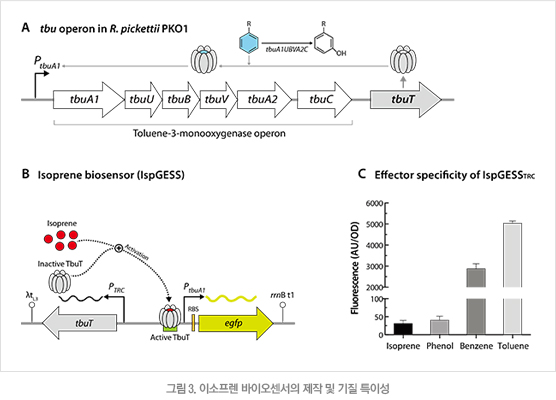
이소프렌에 반응하는 전사 조절 시스템으로 랄스토니아 피케티(Ralstonia pickettii) 유래의 TbuT 전사 인자와 이에 반응하는 PtbuA1 프로모터를 기반으로 하였다. TbuT는 톨루엔(toluene)이 존재할 때 톨루엔 산화효소(toluene-3-monooxygenase)를 생성하는 tbu 오페론을 활성화시키는 단백질이다(그림 3A). 이 전사조절인자는 구조적 유사성에 의하여 이소프렌에 대해서로 영향을 받는 것으로 추정되는 바, 이를 토대로 이소프렌에 의하여 형광이 발현되는 새로운 유전자 회로, IspGESS를 구축하였다(그림 3B). 그 후 톨루엔과 이소프렌을 포함한 다양한 기질에 대한 형광 출력을 측정한 결과 톨루엔에 비해서는 활성이 매우 낮았으나 이소프렌에 대해서도 유의한 수준의 출력을 내는 것으로 확인되었다(그림 3C).
2.2 T7 RNA 중합효소 기반 이소프렌 바이오센서 민감도 개량
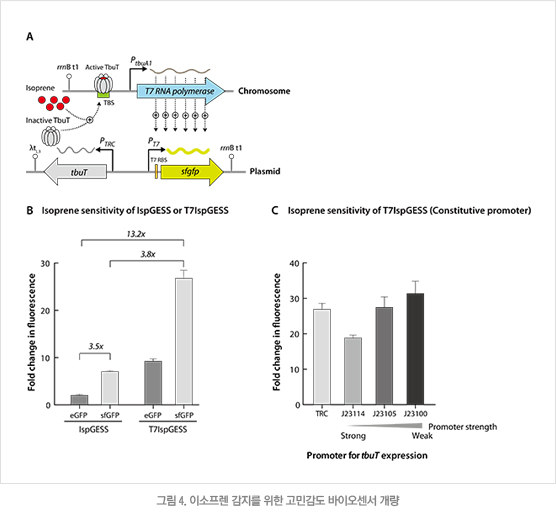
IspGESS를 실재로 활용하기 위해서는 이소프렌에 대한 민감도를 증가시킬 필요가 있었다. 먼저 리포터 단백질을 슈퍼 폴더 녹색 형광 단백질(superfolder GFP, sfGFP)로 교체하였고 이 방법에 의해 이소프렌에 대한 민감도가 3.5배 증가하였다(그림 4B). 두 번째로는 이소프렌이 존재할 때 활성화된 PtbuA1 프로모터가 T7 RNA 중합효소를 발현시키고, 생산된 T7 RNA 중합효소가 형광 단백질의 발현을 가속화하는 연속반응(cascade) 방식의 유전자 회로, T7IspGESS를 제작하였다(그림 4A). 이 방법에 의해 이소프렌에 대한 민감도가 3.8배 증가하여 결과적으로 총 13.2배 민감도가 향상된 이소프렌 바이오센서를 구축할 수 있었다(그림 4B). 또한 전사인자 TbuT의 프로모터를 다양한 세기의 신규 프로모터로 교체하였다. 교체 결과 이소프렌에 대한 민감도는 강한 항시 발현 프로모터인 PJ23100에서 가장 우수하였다(그림 4C).
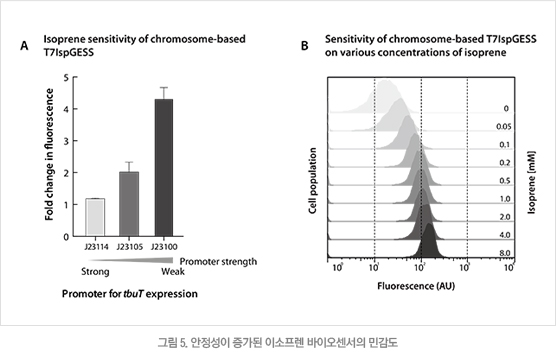
2.3 유전체 내로 이소프렌 유전자 회로 삽입을 통한 바이오센서 안정성 증가
바이오센서의 성능 중 민감도 다음으로 고려되어야 할 사항은 제작된 유전자 회로의 균주 내에서의 안정적 유지이다. 따라서 T7IspGESS 유전자 회로를 대장균의 염색체 내로 삽입하여 이소프렌 바이오센서의 안정성을 증가시켰다.염색체 내에 존재하는 이소프렌 바이오센서의 민감도는 TbuT 단백질의 발현이 높을 때 가장 좋았으며 50μM로 낮은 농도의 이소프렌도 정량 가능하였다(그림 5).
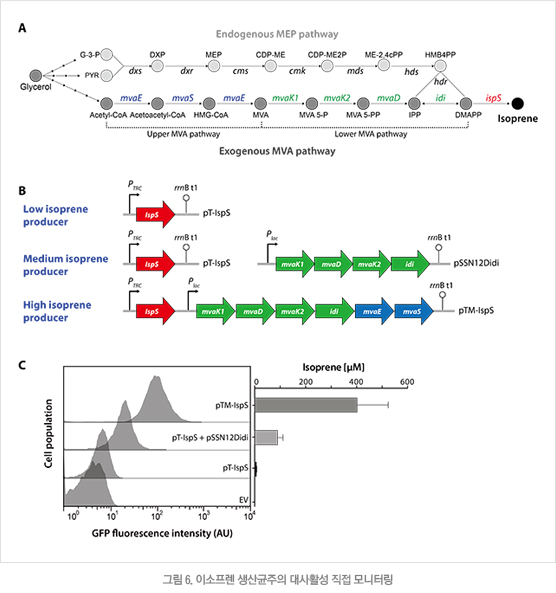
2.4 이소프렌 생산 균주의 활성도 모니터링
이소프렌 바이오센서가 탑재된 유전자 재조합 균주에서 이소프렌 생산을 모니터링하는 실험을 수행하였다. 대장균은 methylerythritol phosphate(MEP) 경로를 통해 이소프렌의 전구체인 DMAPP를 생산할 수 있지만, 외래 효소들로 이루어진 mevalonate(MVA) 경로를 도입할 경우 DMAPP 공급을 크게 증가시킬 수 있다(그림 6A). 따라서 이소프렌 생산량을 다양하게 조절하기 위하여 MVA 경로를 구성하는 효소의 조합이 서로 다른 플라스미드를 각각 제작하였다(그림 6B). 제작된 플라스미드를 대장균에 도입하여 이소프렌을 측정한 결과 5μM부터 400μM까지 다양하게 생산되었다(그림 6C). 가장 낮은 5μM 수준의 이소프렌 생산은 감지하지 못하였으나, 100μM 및 400μM 수준의 이소프렌을 생산에서는 생산량에 비례하여 바이오센서의 형광 출력값이 상승함을 확인하였다(그림 6C).
3. 기대효과
본 연구에서는 낮은 민감도를 갖는 이소프렌 바이오센서를 점진적으로 개량 및 최적화하여 안정적이고 높은 민감도를 갖은 바이오센서로 개량하였다. 또한, 개량된 이소프렌 바이오센서를 이용하여 유전자 재조합 균주에서 생산되는 이소프렌을 정량적으로 감지하는데 성공하였다. 본 기술은 신규한 이소프렌 합성효소(isoprene synthase)를 자연계에서 발굴하거나 이소프렌 합성효소에 돌연변이를 도입하여 활성이 증가된 효소를 선별하는데 활용될 수 있다. 또한 MVA 경로를 구성하는 효소들을 재설계하여 이소프렌을 포함한 다양한 터펜의 생산량을 증가시키는데 기여할 것으로 예측된다.
참고문헌
Choi SL, Rha E, Lee SJ, Kim H, Kwon K, Jeong YS, Rhee YH, Song JJ, Kim HS, and Lee SG (2014). ACS Synthetic Biology, 2014, 3 (3), pp163-171.









 고감도 이소프렌 감지 바이오센서의 개발 및 적용
고감도 이소프렌 감지 바이오센서의 개발 및 적용








