알코올 분해효소 III (Alcohol Dehydrogenase III)의 간섬유화 촉진 기전 규명

한국과학기술원 정원일 교수
Alcohol Dehydrogenase III Exacerbates Liver Fibrosis by Enhancing Stellate Cell Activation and Suppressing Natural Killer Cells in Mice / Hepatology. 2014 Sep;60(3):1044-53.
1. 연구배경
정상적으로 간성상세포들 (hepatic stellate cells; HSCs)은 체내 레티놀 (retinol; vitamin A)의 약 70~80% 정도를 저장하고 있으며 레티놀 항상성(retinol homeostasis)에 중요한 역할은 한다. 일반적으로 레티놀은 여러 가지 효소에 의하여 레티노익산 (retinoic acid)으로 대사되고 핵수용체 (nuclear receptor)인 RARs (retinoic acid receptors)와 RXRs (retinoid X receptors)들과 결합한 후 전사인자로서 다양한 유전자의 발현을 조절하게 된다. 일반적으로 레티놀 대사는 ADHs (alcohol dehydrogenases), SDR (short-chain dehydrogenase/reductase) 및 RDHs (retinol dehydrogenases) 등에 의해 이루어진다고 알려져 있으나 microsome에 위치하는 SDR과 RDH 보다는 세포질에 있는 ADHs가 레티놀 대사에 더 효과적이라고 알려져 있다 있다.
최근 만성 간질환시 레티놀의 간내 함량이 감소하는 반면 레티노익산의 간내 수치는 증가하는 연구결과들이 발표되고 있어, 간질환시 레티놀대사가 증가한다는 것을 암시한다. 현재까지 간성상세포들의 레티놀 저장 및 항상성 역할에 대한 많은 연구가 있었지만, 다양한 간손상시 활성화 되는 간성상세포들에서 어떤 효소에 의해 레티놀 대사가 이루어지는 지 또한 대사된 레티노익산은 어떤 기능을 하는지에 대한 연구는 자세히 알려진 바가 없다. 또한 간섬유화 과정에서 활성화된 간성상세포들 주변에 존재하는 자연살해세포 (natural killer cell; NK cell)들도 간성상세포들이 분비하는 레티놀에 어떤 반응을 보이는지에 대해 알려지지 않은 상태였다 (그림 1). 따라서 이러한 간성상세포의 활성화 혹은 간섬유화과정에 있어서 중요 레티놀 대사 효소가 어떠한 역할을 하는지 밝히고, 간섬유화과정에서 간성상세포와 밀접한 상호작용을 하는 자연살해세포에서의 레티놀 대사의 효과에 대하여 분석하고자 본 연구를 시작하게 되었다.
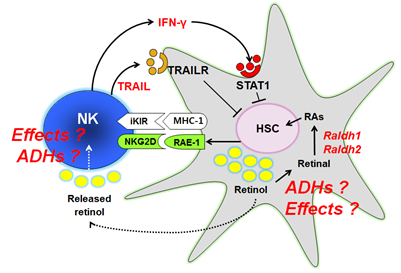
그림 1. 간성상세포와 자연살해세포에서의 레티놀 대사의 역할 규명을 위한 가설
2. 연구결과
먼저 간성상세포에서의 레티놀 대사와 관련되어 가장 중요한 효소를 확인하기 위하여 ADH의 isotype에 대해서 quantitative real-time PCR을 통하여 분석을 시행하였는데, 마우스뿐만 아니라 인체 유래 간성상세포주 (hTERT)에서도 ADH3를 주로 발현한다는 결과를 얻었다 (그림 2). 그리고 4-methylpyrazole을 이용하여 ADH를 억제하여 레티놀 대사를 차단하면 , 레티놀의 양은 증가하고 레티노익산은 감소하게 되는데, 이 경우에 간성상세포의 콜라겐 합성이 상대적으로 억제되는 것을 확인하였다 (그림 3).
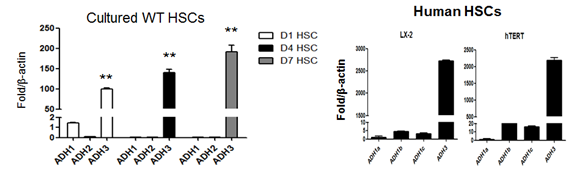
그림 2. 마우스 간성상세포 및 인체 유래 간성상세포주에서의 ADHs 발현 양상
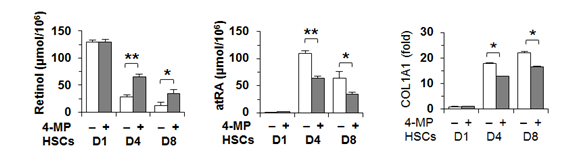
그림 3. 4-methylpyrazole에 의한 ADH 억제 후 간성상세포의 레티놀 및 레티노익산 측정 및 콜라겐 발현 관찰
다음으로 세포실험 결과가 생체에서도 재현되는지를 확인하기 위해서 wild-type (WT)과 ADH3 knockout (ADH3 KO) 마우스에 간섬유화를 유도하였다. ADH3 KO 마우스가 WT 마우스에 비하여 간섬유화가 줄어들어 있었고, 간성상세포의 활성화와 관련된 α-SMA의 발현도 WT에 비하여 상대적으로 억제되어 있었다. 따라서 레티놀 대사가 억제되면 간성상세포의 활성화가 차단되어 결과적으로 간섬유화가 줄어든다는 것을 증명할 수 있었다 (그림 4A). 또한 간섬유화과정에서 자연살해세포의 활성에 미치는 레티놀 대사의 역할을 관찰한 결과, 자연살해세포에서 ADH3가 억제되었을 때 IFN-γ (Interferon gamma) 등의 발현이 증가하는 것을 확인할 수 있었다 (그림 4B). 그리고 in vitro에서 자연살해세포를 간성상세포와 공동배양을 했을 경우에도 ADH3가 결손된 자연살해세포의 경우 IFN-γ 의 분비가 상대적으로 증가하는 것이 관찰되었다 (그림 4C).
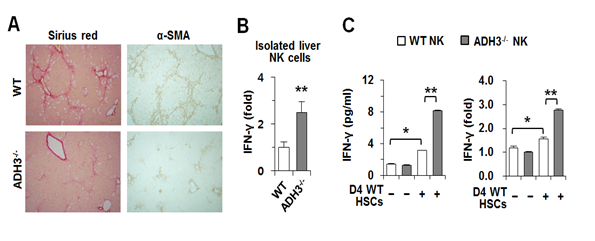
그림 4. WT, ADH3 KO 마우스의 간섬유화 비교 및 자연살해세포에서의 사이토카인 발현 분석
상기에서 관찰된 결과들은 ADH3 유전자가 모든 세포들에서 결손된 마우스들로부터 나온 결과임으로 이를 다시 확인하기 위해, 자연살해세포는 골수 유래세포이지만 간성상세포는 골수 유래세포가 아니기 때문에 간성상세포와 자연살해세포 각각에서 ADH3결손 모델을 만들기 위해 WT과 ADH3 KO 마우스에 WT과 ADH3 KO 골수세포를 각각 주입하여 4종류의 ADH3 chimera (WTWT, WTADH3, ADH3WT, ADH3ADH3) 마우스를 제작하고 chimerism을 확인한 후 간섬유화를 유도하였다 (그림 5A). 이들 마우스에서의 간섬유화의 정도는 WTWT 마우스에서 가장 진행되어 있었고 간성상세포가 ADH3 KO (ADH3WT)이거나 주입한 골수 세포가 ADH3 KO (WTADH3, ADH3ADH3)인 경우에 섬유화의 정도가 감소하였고 간조직에서 α-SMA 발현도 감소한 것을 확인하였다 (그림 5B and C). 또한 ADH3 KO 골수 세포를 주입하였을 때, 자연살해세포(WTADH3, ADH3ADH3)의 IFN-γ 발현이 WTWT에 비하여 상대적으로 증가하였고 이에 따라서 간성상세포의 활성이 억제되어 섬유화가 감소되었다는 것을 증명할 수 있었다 (그림 5D).
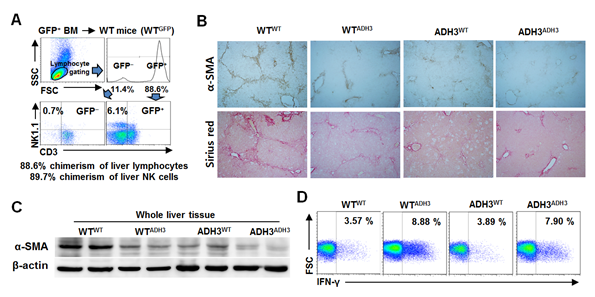
그림 5. ADH3 chimera 마우스를 이용한 간성상세포 및 자연살해세포에서의 ADH3 역할 분석
(위 첨자: 골수 세포 유형; 아래 첨자: 수혜 마우스 유형)
3. 연구의 성과 및 의의
레티놀 대사와 간섬유화 혹은 간성상세포의 활성화의 관계는 확실히 정립되지 않았다. 그러나 본 연구에서는 간성상세포에서 주로 발현되는 레티놀 대사 효소가 ADH3라는 것을 밝혔고, 이를 억제하면 간성상세포의 활성화가 줄어들고 결과적으로 간섬유화의 진행도 억제할 수 있다는 것을 세포 실험 및 마우스 생체 실험을 통해서 증명하였다. 또한 ADH3 KO 마우스의 경우에는 간섬유화 과정에서 자연살해세포의 IFN-γ 발현이 증가하여 간성상세포를 억제하고 나아가 간섬유화의 진행을 억제한다는 결과를 확인할 수 있었다. 따라서 본 연구는 ADH3는 간성상세포와 자연살해세포에서 각각 세포에 특이적인 작용을 통하여 간섬유화의 발전을 촉진하는 효소라는 것을 확인함으로써 ADH3가 간섬유화에 대한 새로운 치료 타겟이 될 수 있음을 보여주었다 (그림 6A and B). 본 연구를 바탕으로 현재까지 치료약이 없었던 간섬유화 및 간경변증에 대한 신약개발이 촉진될 것으로 사료된다.
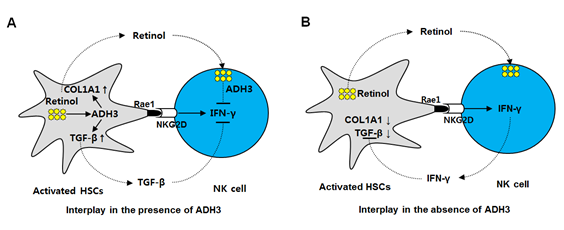
그림 6. 간섬유화과정에서 간성상세포와 자연살해세포의 활성에 대한 ADH3의 세포 특이적인 역할
참고문헌
1. Friedman SL. Hepatic stellate cells: protean, multifunctional, and enigmatic cells of the liver. Physiol Rev 2008;88:125-172.
2. Pares X, Farres J, Kedishvili N, Duester G. Medium- and short-chain dehydrogenase / reductase gene and protein families: medium-chain and short-chain dehydrogenases/reductases in retinoid metabolism. Cell Mol Life Sci 2008;65:3936-3949.
3. Duester G, Mic FA, Molotkov A. Cytosolic retinoid dehydrogenases govern ubiquitous metabolism of retinol to retinaldehyde followed by tissue-specific metabolism to retinoic acid. Chem Biol Interact 2003; 143-144:201-210.
4. Natarajan SK, Thomas S, Ramachandran A, Pulimood AB, & Balasubramanian KA (2005) Retinoid metabolism during development of liver cirrhosis. Arch Biochem Biophys 443(1-2):93-100.
5. Leo MA & Lieber CS (1982) Hepatic vitamin A depletion in alcoholic liver injury. N Engl J Med 307(10):597-601.
6. Jeong WI, Park O, Suh YG, Byun JS, Park SY, Choi E, et al. Suppression of innate immunity (natural killer cell/interferon-gamma) in the advanced stages of liver fibrosis in mice. HEPATOLOGY 2011;53:1342-1351.









 알코올 분해효소 III (Alcohol Dehydrogenase III)의 간섬유화 촉진 기전 규명
알코올 분해효소 III (Alcohol Dehydrogenase III)의 간섬유화 촉진 기전 규명








