진세노사이드 전구체 합성생물학 연구 동향 및 추세 보고서
최보현, 이평천
아주대학교
1. 개요
전 세계적으로 기후변화나 환경에 영향을 주지 않는 에너지와 합성 화학 물질의 공급요구를 대체할 수 있는 합성 생물학을 이용한 재사용 에너지와 화학물질의 발전이 이루어지고 있다.아이소프레노이드 (Isoprenoids)는 식품, 의학, 약학, 바이오 연료 및 화학 등 다양한 분야의 산업체에 적용되고 있는 유기 합성체의 큰 그룹이다. 이러한 다양한 구조의 아이소프레노이드는 생합성 전구체인 Isopentenyl diphosphate (IPP)와 이성질체인 dimethylallyl diphosphate (DMAPP)로부터 합성되며, 이러한 전구체는 mevalonate (MVA) pathway와 2-C-methly-D-erythritol 4-diphosphate (MEP) pathway 두 개의 독립적인 생합성 대사회로를 통해 생산된다.(그림 1.)
Isoprenoids 계열 물질들은 보통 식물체로부터 분리되며 이를 이용해 anticancer drugs, antimalarial drugs, 상업적 용도의 flavor, fragrance등에 사용되고 있다. 그러나 식물체로부터의 조직 추출을 통한 분리는 일반적으로 낮은 수율의 양을 얻을 수 있어 microbial host를 이용한 높은 수율의 product를 얻는 연구의 본격화가 절실한 시점이다. 본 보고서에서는 이런 Isoprenoids 전구체의 합성생물학적 연구 동향을 알아보고 이를 이용한 현시점에서의 발전 방향성을 고찰하고자 한다.
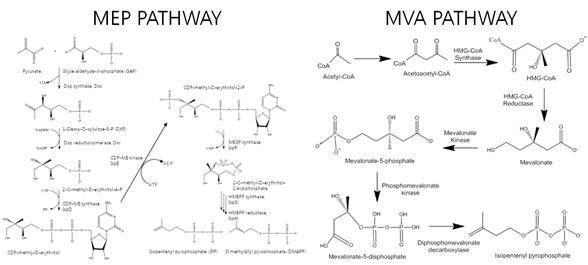
그림 1. Isoprenoids의 전구체 생합성회로: MEP 대사회로와 MVA 대사회로
(Jonathan Lombard & David Moreira, 2011)
2. Isoprenoids 전구체 대사회로 유전자의 다양성
일반적으로 Listeria monocytogenes 와 특정 streptomyces 종과 같이 두 개의 IPP 생합성 대사회로를 모두 가진 경우(Boucher & Doolittle et al, 20000)를 제외하고는 동물계에서 IPP 생합성 과정은 MVA 대사회로에, 고세균은 약간의 변형된 MVA대사회로에 특화되어 있으며 대부분의 박테리아 종의 경우 두 개중 하나의 대사회로가 독점적으로 존재 한다. MVA 대사회로에는 총 6개의 효소가 관여하며 이 효소들은 acetyl-CoA 와 acetoacetyl-CoA를 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA)로 전환하며 중간 대사체인 mevalonate에 phosphorylation 과정을 통해 phosphomevalonate와 diphosphomevalonate를 합성하며 최종적으로 C5 building block인 IPP나 DMAPP로 전환시킨다.(그림 1.)
MVP에 비해 MEP 대사회로의 경우 8 단계의 효소 활성 과정을 필요로 하며 pyruvate와 glyceraldehyde 3-phosphoate를 1-deoxy-D-xylulose 5-phosphate (DOXP)로 합성하는 첫 번째 단계를 거쳐 DXR에 의해 2C-methyl-D-erythritol 4-phosphate (MEP)로 전환된다. 4-diphosphocytidyl-2c-methyl-D-erythritol가 IspD의 촉매로 인해 CTP와 MEP 반응으로 합성된 후 IspE에 의해 CDP-ME2P로 합성되고 마지막으로 IspF, IspG, IspH에 의해 IPP로 합성된다. (그림 1.)
이러한 Isoprenoids 생합성 전구체인 IPP의 생합성 대사회로에 대한 연구는 다양한 미생물에 대하여 집중적으로 진행되어 왔다. 최근에는 phylogenetic 연구를 통한 MVA 대사회로 유전자들의 유래 및 종류에 대한 연구가 활발히 진행되고 있으며 이를 통해 MEP 대사회로와 MVA 대사회로의 진화론적 연구가 진행 중이며, 다양한 미생물 종들의 MVA 대사회로 유전자 시스템이 밝혀지고 있는 추세이다.( 표 1.)
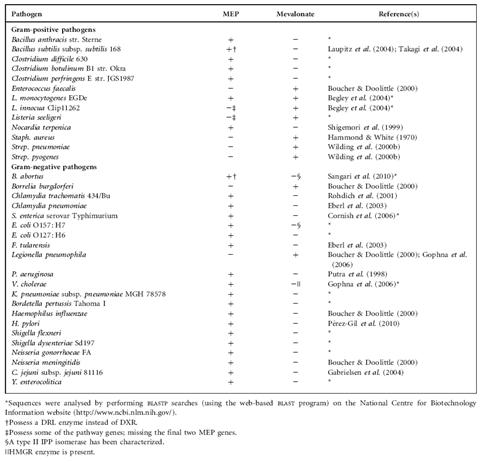
표 1. Archaeal Genera와 Bacteria에 관계된 MVA 대사회로 유전자들의 종별 비교
(Jonathan Lombard & David Moreira, 2011)
3. 진세노사이드 전구체 대사회로의 진화론적 방향성
생물계의 세 개의 큰 도메인 (원핵생물계, 진핵생물계, 고생물계)에서 진세노사이드 전구체인 IPP의 생합성 대사 경로는 매우 복잡하지만 MEP 대사회로는 박테리아에서, 일반적인 MVA 대사회로는 진핵생물계, 변형된 MVA 대사회로는 고생물계라는 명확한 관계가 있다. phylogenetic 연구를 통한 가설을 보면 MVA 와 MEP 대사회로의 진화론적 시나리오 3가지 가능성이 있다는 주장이 있다.(Lombard and Moreira, 2010). 첫 번째 가설은 Isoprenoids 전구체 대사회로가 존재하지 않던 초기 조상으로부터 3개의 도메인으로 진화과정에서 각각의 전구체 대사회로가 나타났다는 설(그림 2.A)이며, 두 번째 가설은 초기 조상에 MEP, MVA, MVA-Arc 대사회로가 모두 존재한 상태에서 3개 도메인으로 진화과정에서 각 대사회로에 맞게 특화되었다는 설이다(그림 2.B ). 마지막 세 번째 가설은 초기 조상에서 MVA 대사회로를 가진 상태에서 3개 도메인으로 진화 과정에서 각 대사회로에 맞게 변이가 일어났다는 설이다(그림 2.C).
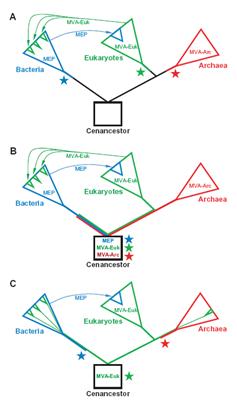
그림 2. 생물학적 세 개 도메인에서의 다른 Isoprenoid 생합성 대사회로의 예상 진화도.
MEP 대사회로는 파란색, 일반 MVA 대사회로는 초록색, 변형 MVA 대사회로는
붉은색으로 표시되었음. 별모양은 각 대사회로의 기원을 나타내었음.
4. 합성생물학적 적용 기술 동향
최근 환경에 미치는 영향이 크던 기존의 합성 기술의 보완을 위해 친환경 기술의 발전이 장려되는 추세에서 합성생물학을 통한 여러 연구가 이루어지고 있다. 진세노사이드를 포함한 고기능성 Isoprenoids에 대한 가치의 증가로 의학, 약학 분야에서 주로 연구가 이루어지고 있으며 산업화를 위한 생산량/수율 증대의 목적으로 전구체인 IPP 대사회로의 연구 또한 범위가 확대되어가는 추세이다.
MEP 대사회로의 전체 흐름에 대한 완전한 정보가 최근에 밝혀진 이래로 MEP 대사회로를 통한 연구가 활발히 진행되고 있으며 주로 의학적 분야에서 MEP 대사회로의 핵심이 되는 효소의 발현을 억제하는 inhibitor를 이용한 항박테리아 물질의 개발 (Handa. S et al, 2013, Guobin Cai et al. 2013) 및 MEP 대사회로에 관여하는 효소 자체에 mutation engineering 기법을 적용한 재조합 효소를 이용해 인체에 손상 조직에서 발생하는 효과에 따른 약학적 연구 (Parul Gupta et al 2013) 역시 진행되고 있다.
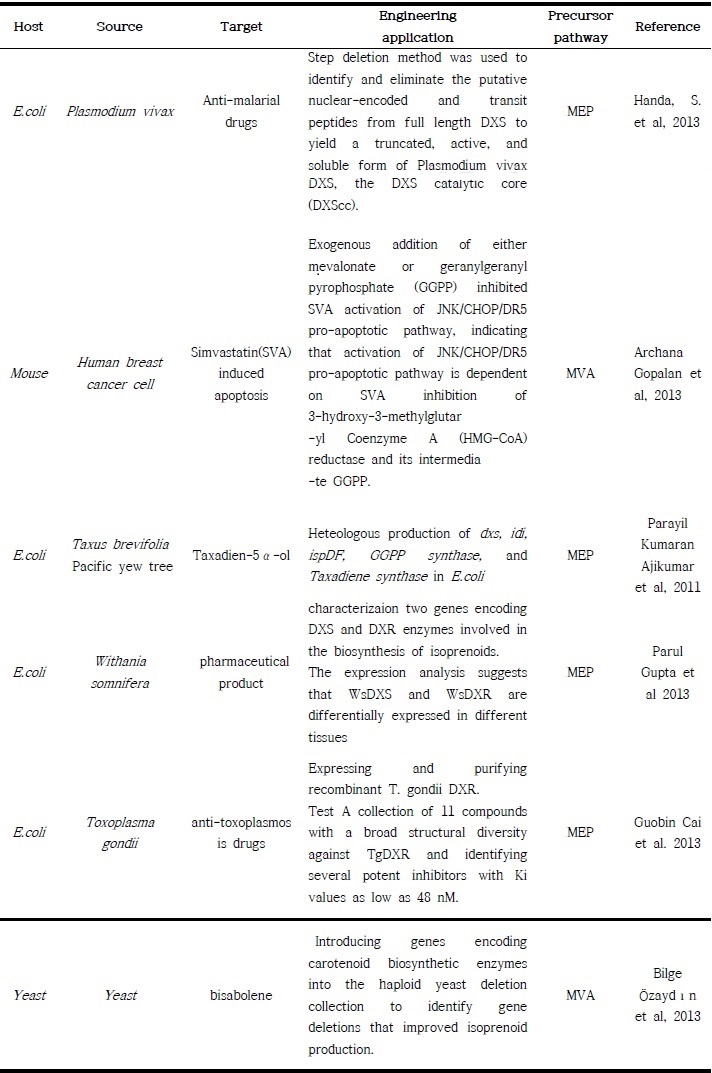
MVA 대사회로의 경우 인체 내 sterol 계통에 관여하는 중요한 대사회로이기 때문에 의학적, 약학적 분야에서 많은 연구가 진행되고 있으며 최근에는 산업적인 용도로의 적용을 위한 연구 역시 활발하게 진행되고 있다. 의학적, 약학적 분야에서는 MEP 대사회로에서의 연구와 마찬가지로 대사과정에서 핵심 역할을 하는 효소의 활성을 억제하는 (주로 3-hydroxy-3-methylglutaryl-CoA reductase) Simvastatin과 같은 inhibitor를 이용한 cancer cell의 apoptosis를 유발시키는 연구 (Archana Gopalan et al, 2013)나 IPP와 DMAPP의 과발현을 통한 유용 Isoprenoid 물질의 생산량/수율 증대를 위한 연구 또한 활발히 진행 중이다. 또는 대사회로에 존재하는 여러 유전자들 중 불필요한 유전자의 제거를 통한 대사물질의 과발현 시스템을 구축하는 연구 (Bilge Özaydın et al, 2013) 등 다양한 approach로 Isoprenoid 전구체 대사회로에 대한 합성생물학적 연구가 이루어지고 있는 추세이다.(표 3.)
그러나 이러한 IPP 전구체 대사회로의 광범위한 적용 범위와 잠재력에 비해 연구는 아직 초기화 단계이며 활성화 또한 미미한 실정이다. 따라서 고부가 물질인 진세노사이드를 대량생산할 수 있도록 IPP 전구체 대사회로에 대한 기존의 낮은 수율의 제약을 뛰어넘을 수 있는 IPP 전구체 과발현 시스템의 구축이 합성생물학을 토대로 이루어져야 한다.
표 3. IPP 생산 MEP와 MVA 대사회로를 이용한 다양한 합성생물학적 적용 연구
5. 참고자료
1. Ajikumar, P.K, Xiao, W.-H, Tyo, K.E.J, Wang, Y, Simeon, F, Leonard, E, Mucha, O, Phon, T.H, Pfeifer, B , Stephanopoulos, G., Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli, 2010, Science, 330, 70-74
2. Archana Gopalan, Weiping Yu, Bob G. Sanders, Kimberly Kline, Simvastatin inhibition of mevalonate pathway induces apoptosis in human breast cancer cells via activation of JNK/CHOP/DR5 signaling pathway, 2013, cancer letter, 329, 9-16
3. Begley, M., Bron, P. A., Heuston, S., Casey, P. G., Englert, N.,Wiesner, J., Jomaa, H., Gahan, C. G. M. & Hill, C., Analysis of the isoprenoid biosynthesis pathways in Listeria monocytogenes revealsa role for the alternative 2-C-methyl-D-erythritol 4-phosphate pathway in murine infection. 2008, Infect Immun 76, 5392–5401.
4. Boucher, Y. & Doolittle, W. F., The role of lateral gene transfer in the evolution of isoprenoid biosynthesis pathways. 2000, Mol Microbiol 37, 703-716.
5. Cornish, R. M., Roth, J. R. & Poulter, C. D., Lethal mutations in the isoprenoid pathway of Salmonella enterica. 2006, J Bacteriol 188, 1444–1450.
6. Eberl, M., Hintz, M., Reichenberg, A., Kollas, A. K., Wiesner, J. &Jomaa, H.. Microbial isoprenoid biosynthesis and human cd T cell activation. 2003, FEBS Lett 544, 4–10.
7. Gabrielsen, M., Rohdich, F., Eisenreich, W., Gra¨wert, T., Hecht, S.,Bacher, A. & Hunter, W. N.. Biosynthesis of isoprenoids: a bifunctional IspDF enzyme from Campylobacter jejuni. 2004, Eur J Biochem 271, 3028–3035.
8. Gophna, U., Thompson, J. R., Boucher, Y. & Doolittle, W. F., Complex histories of genes encoding 3-hydroxy-3-methylglutarylCoenzymeA reductase. 2006, Mol Biol Evol 23, 168–178.
9. Hammond, R. K. & White, D. C., Carotenoid formation by Staphylococcus aureus. 1970, J Bacteriol 103, 191–198.
10. Handa, S.a, Ramamoorthy, D, Spradling, T.J. Guida, W.C, Adams, J.H, Bendinskas, K.G, Merkler, D.J, Production of recombinant 1-deoxy-d-xylulose-5-phosphate synthase from Plasmodium vivax in Escherichia coli, 2013, FEBS Open Bio, 3, 124-129
11. Jonathan Lombard and David Moreira, origins and Early Evolution of the Mevalonate Pathway of Isoprenoid Biosynthesis in the Three Domain of Life, 2011, Mol. Biol, Evol, 28(1), 87-99.
12. Laupitz, R., Hecht, S., Amslinger, S., Zepeck, F., Kaiser, J., Richter,G., Schramek, N., Steinbacher, S., Huber, R. & other authors., Biochemical characterization of Bacillus subtilis type II isopentenyl diphosphate isomerase, and phylogenetic distribution of isoprenoid biosynthesis pathways. 2004, Eur J Biochem 271, 2658–2669.
13. Parul Gupta, Aditya Vikram Agarwal, Nehal Akhtar, Rajender Singh Sangwan, Surya Pratap Singh, Prabodh Kumar Trivedi, Cloning and characterization of 2-C-methyl-d-erythritol-4-phosphate pathway genes for isoprenoid biosynthesis from Indian ginseng, Withania somnifera, 2013, Protoplasma, 250, 285-295
14. Pe´rez-Gil, J., Bergua, M., Boronat, A. & Imperial, S., Cloning and functional characterization of an enzyme from Helicobacter pylori that catalyzes two steps of the methylerythritol phosphate pathway for isoprenoid biosynthesis. 2010, Biochim Biophys Acta 1800, 919–928.
15. Puan, K. J., Wang, H., Dairi, T., Kuzuyama, T. & Morita, C. T., fldA is an essential gene required in the 2-C-methyl-D-erythritol 4-phosphate pathway for isoprenoid biosynthesis. 2005, FEBS Lett 579, 3802–3806.
16. Rohdich, F., Wungsintaweekul, J., Fellermeier, M., Sagner, S., Herz, S., Kis, K., Eisenreich, W., Bacher, A. & Zenk, M. H., Cytidine 59-triphosphate-dependent biosynthesis of isoprenoids: YgbP protein of Escherichia coli catalyzes the formation of 4-diphosphocytidyl-2-Cmethylerythritol. 1999, Proc Natl Acad Sci U S A 96, 11758–11763.
17. Sangari, F. J., Pe´rez-Gil, J., Carretero-Paulet, L., Garcı´a-Lobo, J. M. & Rodrı´guez-Concepcio´ n, M. A new family of enzymes catalyzing the first committed step of the methylerythritol 4-phosphate (MEP) pathway for isoprenoid biosynthesis in bacteria. 2010, Proc Natl Acad Sci U S A 107, 14081–14086.
18. Sinead Heuston, Maire Begley, Cormac G.M. Gahan and Colin Hill, Isoprenoid Biosynthesis in Bacterial Pathogens, 2012, Microbiology 158, 1389-1401.
19. Takagi, M., Kaneda, K., Shimizu, T., Hayakawa, Y., Seto, H. & Kuzuyama, T. Bacillus subtilis ypgA gene is fni, a nonessential gene encoding type 2 isopentenyl diphosphate isomerase. 2004, Biosci Biotechnol Biochem 68, 132–137
20. Wilding, E. I., Kim, D.-Y., Bryant, A. P., Gwynn, M. N., Lunsford, R. D., McDevitt, D., Myers, J. E., Jr, Rosenberg, M., Sylvester, D. & other authors . Essentiality, expression, and characterization of the class II 3-hydroxy-3-methylglutaryl coenzyme A reductase of Staphylococcus aureus. 2000, J Bacteriol 182, 5147–5152.









 진세노사이드 전구체 합성생물학 연구 동향 및 추세 보고서(아주대 최보현, 이평천)
진세노사이드 전구체 합성생물학 연구 동향 및 추세 보고서(아주대 최보현, 이평천)






